

§5.10. Межмолекулярный перенос энергии в биоструктурах
5.10.1. Химический и физический способы переноса энергии
Структурно-динамическая организация макромолекул непосредственно связана с элементарными актами функциональных процессов, в которых участвуют макромолекулы. К элементарным актам можно отнести перенос массы (см., например, перенос лигандов, с. 489–490) и процессы, связанные с трансформацией электронной энергии (изменением электронного состояния) макромолекул: перенос электрона (см. §5.11, с. 578) и перенос электронного возбуждения (§5.10), не сопровождающийся переносом электрона.
Перенос энергии электронного возбуждения, наряду с переносом электронов, играет важную роль в биологических процессах. Он может осуществляться двумя путями: химическим процессом или физическим.
Химический способ передачи
связан с химическими превращениями,
образованием промежуточных соединений
(см., например, с. 509-510). В этом случае
избыток электронной энергии сосредоточен
на связях в нестабильных промежуточных
соединениях. Чтобы энергия связей не
успела перейти в тепловую энергию, весь
процесс трансформации энергии должен
происходить за время, не превышающее
характерное время
![]() с
рассеяния энергии при
электронно-колебательной релаксации.
Последовательное осуществление
процессов, обеспечивающих передачу
энергии, за столь короткое время
представляет собой значительные
трудности. Кроме того, поскольку в
процессах передачи энергии принимают
участие низкомолекулярные соединения,
разделенные пространственно и
диффундирующие к месту реакции, то
химические процессы передачи и
трансформации энергии должны происходить
в специальных комплексах.
с
рассеяния энергии при
электронно-колебательной релаксации.
Последовательное осуществление
процессов, обеспечивающих передачу
энергии, за столь короткое время
представляет собой значительные
трудности. Кроме того, поскольку в
процессах передачи энергии принимают
участие низкомолекулярные соединения,
разделенные пространственно и
диффундирующие к месту реакции, то
химические процессы передачи и
трансформации энергии должны происходить
в специальных комплексах.
Физический способ, не
сопровождающийся ни химическим изменением
вещества, ни переносом каких-либо частиц
или квантов света, представляет собой
миграцию энергии, то есть
безызлучательный обмен энергией
между электронно-возбужденной молекулой
![]() (донором) и молекулой
(донором) и молекулой
![]() ,
находящейся в основном энергетическом
состоянии (акцептором):
,
находящейся в основном энергетическом
состоянии (акцептором):
![]() . (5.23)
. (5.23)
Миграция может происходить, как между разными молекулами, так и одинаковыми, а также внутри одной макромолекулы, но всегда перенос энергии осуществляется на более низкий или такой же энергетический уровень. Физический способ передачи энергии является основным в биологических объектах.
Важную роль в биологических процессах играют процессы переноса энергии фотовозбуждения молекул. Эта энергия, с одной стороны, может перерабатываться в энергию химических связей и в такой форме запасаться в организме, а, с другой стороны, может расходоваться на преодоление активационных барьеров на начальной стадии реакции, то есть стимулировать химические реакции в реакционных центрах.
Вопрос о том «Как это происходит, каким образом энергия управляет жизнедеятельностью? Как она приводит в движение живую машину?» — является одним из кардинальных вопросов биоэнергетики.
5.10.2. Фотовозбуждение сложных молекул
Макромолекулы обладают огромным числом различных (колебательных и вращательных) степеней свободы составляющих ее атомов и атомных групп. Наличие этих степеней свободы и связанная с ними энергия приводят к расщеплению основных энергетических уровней молекулы в полосы, состоящие из близко расположенных дискретных уровней (рис. 5–19).
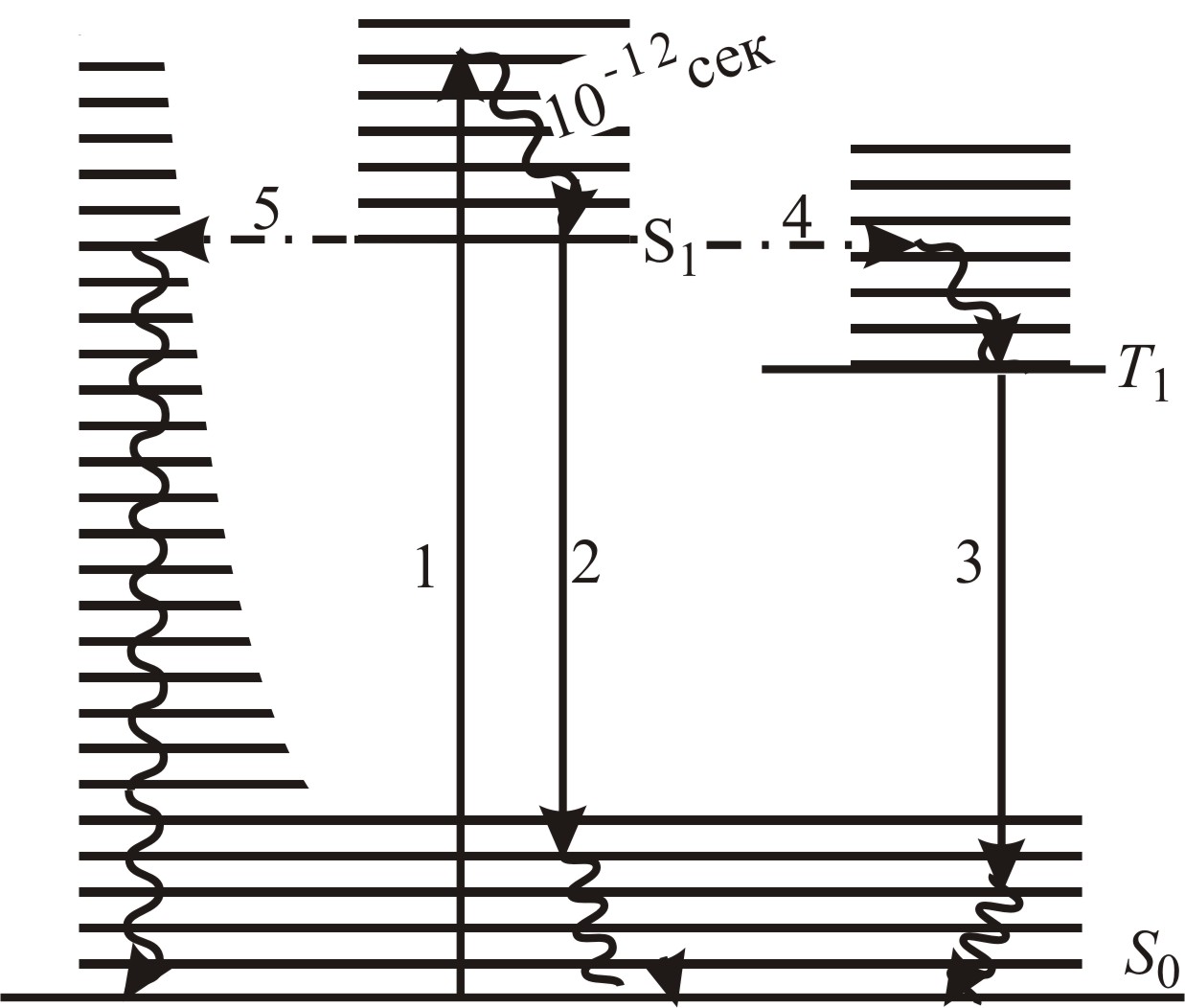
Рис. 5–19. Схема квантовых переходов при молекулярной люминесценции. S0 – основной электронный уровень (сверху – колебательные уровни); возбужденные электронные уровни энергии: синглетный – S1 и триплетный (метастабильный) – T1. Прямые вертикальные стрелки обозначают: 1 – процесс поглощения; 2 – излучательные переходы: флуоресценции (быстрозатухающей люминесценции) и 3 – фосфоресценции (длительной люминесценции); горизонтальные штрихпунктирные стрелки – безызлучательные переходы: интеркомбинационная конверсия (4) и внутренняя конверсия (5), приводящая к безызлучательному переходу энергии возбуждения в тепловую энергию через колебательные степени свободы; волнистые стрелки – процессы колебательной релаксации энергии возбуждения с характерным временем ~(10–1110–13) c, значительно меньшим времени жизни возбужденного электронного состояния
Предположим, что в основном (невозбужденном) состоянии все π-состояния на уровне S0 заполнены электронами, а уровень S1 свободен, то есть исходное состояние является синглетным с суммарным спином электронов, равным нулю. При поглощении кванта света один из π-электронов на уровне S0 перебрасывается в полосу S1 с сохранением спина, так как спин фотона равен нулю. Избыток, по сравнению с энергией уровня S1, энергии электрона расходуется на возбуждение колебаний (волнистая стрелка), соответствующих различным степеням свободы молекулы. При этом электрон, сохраняя ориентацию спина, теряет кинетическую энергию и переходит за время (электрон-колебательной релаксации) t » » (10–11÷10–13) с на нижний уровень полосы S1. Время жизни t* возбужденного синглетного состояния молекулы на этом уровне составляет обычно
![]() . (5.24)
. (5.24)
Далее могут реализоваться четыре возможности.
Во-первых, через время t*
электрон может вернуться на уровень S0
с испусканием кванта света люминесценции
с частотой
![]() .
Люминесценция (от лат. lumenis
– свет, -escent – суффикс,
означающий слабое действие) – это
свечение вещества (избыток над тепловым
излучением), возникающее после поглощения
им энергии возбуждения, причем время
свечения значительно превышает период
световой волны. В природе примерами
люминесценции являются северное сияние,
свечение некоторых минералов, гниющего
дерева и др.
.
Люминесценция (от лат. lumenis
– свет, -escent – суффикс,
означающий слабое действие) – это
свечение вещества (избыток над тепловым
излучением), возникающее после поглощения
им энергии возбуждения, причем время
свечения значительно превышает период
световой волны. В природе примерами
люминесценции являются северное сияние,
свечение некоторых минералов, гниющего
дерева и др.
Во-вторых, возможен безызлучательный переход (5 на рис. 5–19) в исходное состояние. Безызлучательные квантовые переходы происходят между нестационарными состояниями квантовой системы. Энергия релаксирующего в основное состояние электрона расходуется на возбуждение различных степеней свободы ядерной системы, а также квазичастичных возбуждений: фононов, экситонов и др.
В-третьих, поскольку электрон
находится на нижнем уровне S1
значительно большее время, чем время,
в течение которого он релаксировал на
него после возбуждения (![]() )
(2), то за это время в результате рассеяния
на фононах его спин может изменить
направление на противоположное. При
этом электрон переходит на уровень Т
(4 на рис. 5–19), а молекула в триплетное
состояние с параллельно ориентированными
спинами на уровнях S0
и Т. Теперь, чтобы электрон мог
перейти на уровень S0,
и молекула вернулась в синглетное
состояние, спин электрона должен изменить
ориентацию. Вероятность этого процесса
мала и, соответственно, время жизни
электрона на уровне Т значительно
больше, чем на уровне S1
и составляет
)
(2), то за это время в результате рассеяния
на фононах его спин может изменить
направление на противоположное. При
этом электрон переходит на уровень Т
(4 на рис. 5–19), а молекула в триплетное
состояние с параллельно ориентированными
спинами на уровнях S0
и Т. Теперь, чтобы электрон мог
перейти на уровень S0,
и молекула вернулась в синглетное
состояние, спин электрона должен изменить
ориентацию. Вероятность этого процесса
мала и, соответственно, время жизни
электрона на уровне Т значительно
больше, чем на уровне S1
и составляет
tТ » (10–6÷10–2) с. (5.25)
Переходы (3) с уровня Т на S0 практически всегда сопровождаются излучением.
В четвертых, при наличии взаимодействия данной молекулы с окружающими ее молекулами, существует вероятность передачи возбуждения соседям путем резонансного безызлучательного взаимодействия.
Механизмы переноса энергии от возбужденной молекулы к соседней определяются типом их взаимодействия.
Известны несколько механизмов миграции энергии: индуктивно-резонансный, экситонный, обменно-резонансный и полупроводниковый (зонная проводимость).
Основные закономерности переноса энергии электронного возбуждения были открыты при исследовании оптических и фотоэлектрических свойств молекулярных жидкостей (§5.8, с. 236), молекулярных кристаллов (т.1, с.253) и кристаллов, активированных редкоземельными ионами.