

Часть 1. ОБМЕН УГЛЕВОДОВ
Следует добавить:
Виды метаболизма глюкозы – сорбитол, глукуронат, фосфоилирование.
Схему метаболизма г-6-Ф.
Углеводы - это полиоксикарбонильные соединения и их производные. Основными углеводами организма человека являются:
1. Моносахариды (глицеральдегид, диоксиацетон, эритроза, рибоза, дезоксирибоза, рибулоза, ксилулоза, глюкоза, галактоза, фруктоза, манноза, арабиноза, фукоза, и др.);
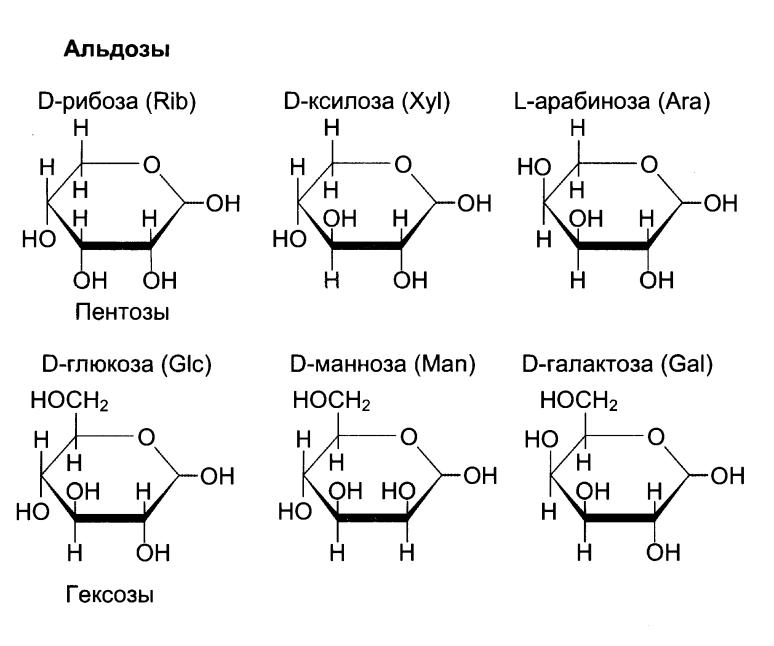

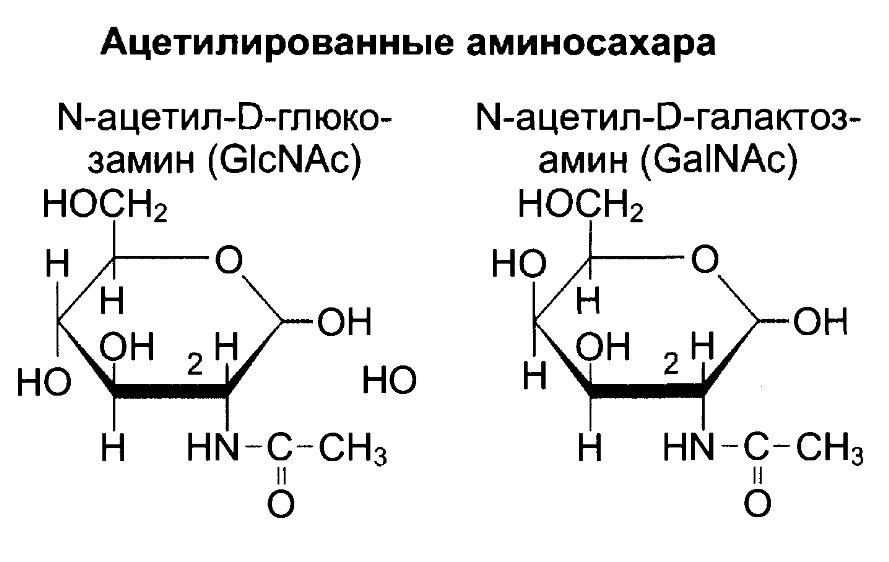


Фукоза – 6-дезоксигалактоза.
α(1→3) связывание с фукозой представляет углеводный антиген IgE-зависимой аллергии.
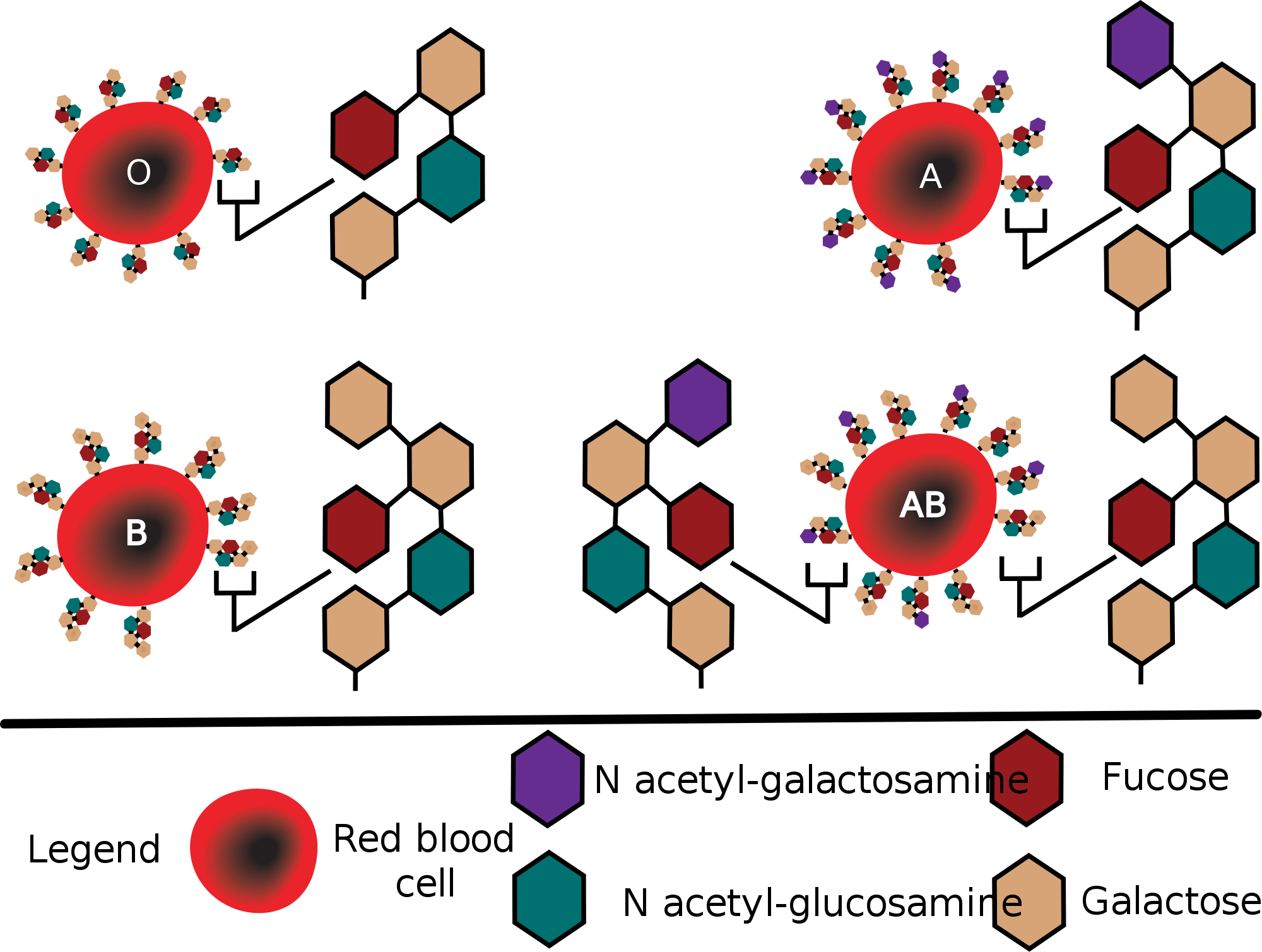
связанная α-1,2- гликозидной связью с галактозой образует H антиген, структуру антигенов групп крови А и В.
При фагоцитозе бактерий или старых клеток организма хозяина происходит распознавание концевых сахаридных групп — глюкозы, галактозы, фукозы, маннозы и др., которые представлены на поверхности фагоцитируемых клеток. Распознавание осуществляется лектиноподобными рецепторами соответствующей специфичности, в первую очередь маннозосвязывающим белком и селектинами, присутствующими на поверхности фагоцитов.
Накопление фукозы в поверхностных гликопротеидах свидетельствует о канцерогенезе и малигнизации. Нпример в составе антигенов Lewis. Например, CD15 (3-фукозил-N-ацетиллактозамин, Lewis X) — углеводная молекула адгезии, опосредует фагоцитоз и хемотаксис нейтрофилов. Интенсивная экспрессия CD15 наблюдается при болезни Ходжкина, при некоторых хронических B-лимфобластомных лейкемиях и в большинстве случаев острой нелимфобластомной лейкемии. CD15 является маркёром плюрипотентных стволовых клеток, принимает участие в адгезии и миграции клеток в эмбрионе.

Сорбитол - шестиатомный спирт, обладающий сладким вкусом. Получают путём гидрирования глюкозы с заменой альдегидной группы на гидроксильную. Используется в производстве аскорбиновой кислоты. Зарегистрирован в качестве пищевой добавки как E420.
Сорбит часто применяется как заменитель сахара, его можно встретить в диетических продуктах и диетических напитках (например, жевательной резинке без сахара). В естественном виде встречается в косточковых плодах, водорослях, высших растениях.
2. Олигосахариды:
Лактоза – молочный сахар, важнейший дисахарид молока млекопитающих. В коровьем молоке – 5%, в женском – 8%. Состоит из галактозы и глюкозы, соединённых β-1,6-гликозидной связью.
Мальтоза – дисахарид, образуется при расщеплении крахмала в кишечнике, либо поступает с продуктами имеющими частично гидролизованный крахмал (солод, пиво). Состоит из двух остатков глюкозы, соединённых α-1,4-гликозидной связью. Изомальтоза - α-1,6-гликозидной связью.
Сахароза – относительно новый дисахарид в питании человека, состоит из α-D-глюкозы и β-D-фруктозы, соединённых α-1,2-гликозидной связью. Так как обе аномерные ОН-группы остатков глюкозы и фруктозы участвуют в образовании гликозидной связи – сахароза не относится к восстанавливающим сахарам. Имеет сладкий вкус. Источник – растения, особенно сахарная свекла и сахарный тростник.
3. Полисахариды. По составу входящих мономеров делятся на:
Гомополисахариды
крахмал, состоит из сотен тысяч остатков глюкозы (гомогликан). Представлен в виде амилозы и амилопектина:
амилоза – неразветвлённый полисахарид из 200-300 остатков глюкозы, соединённых α-1,4-гликозидной связью. Имеет вид спирали. Синий цвет при окраске йодом – следствие наличия этой спирали.
Амилопектин – имеет разветвлённую структуру (α-1,6-гликозидная связь) линейные участки по 20-25 остатков глюкозы.
гликоген – полисахарид животных и человека. Имеет более выраженную частоту ветвлений, чем крахмал (~ через 10 мономеров). Выполняет резервную функцию поддержания строго определённого уровня глюкозы в организме. Не имеет существенного пищевого значения.
Клетчатка (целлюлоза) – основной структурный полисахарид растений. Самое распространённое органическое соединение на земле. Его доля в растениях составляет 40-50%, м.в. – до 103 kDa. Линейный полисахарид, гомогликан, построен из остатков глюкозы, соединённых β-1,4-гликозидной связью. У человека нет ферментов гидролизующих β-1,4 связи, поэтому целлюлоза неиспользуемый углевод.
Пектин - пекти́новые вещества́ (от др.-греч. свернувшийся, замёрзший) — полисахариды, образованные остатками главным образом галактуроновой кислоты. Присутствуют во всех высших растениях, особенно много во фруктах и в некоторых водорослях. Способствуют поддержанию в растениях тургора, повышают засухоустойчивость и устойчивость овощей и фруктов при хранении. Комплексообразующая способность основана на взаимодействии молекулы пектина с ионами тяжелых и радиоактивных металлов. Специальные препараты, включают в рацион питания лиц, находящихся в среде, загрязненной радионуклидами, и имеющих контакт с тяжелыми металлами. Специальные высокоочищенные пектины могут быть отнесены к незаменимому веществу для использования в производстве функциональных пищевых продуктов, а также продуктов здорового и специального (профилактического и лечебного) питания. Оптимальная профилактическая дозировка специального пектина составляет 5-8 г в сутки, а в условиях радиоактивного загрязнения — не менее 15-16 г.
Гетерополисариды (гиалуроновая кислота, хондроитинсульфат, дерматансульфат, кератансульфат, гепарин).
По выполняемым функциям можно разделить на три основные группы:
Резервные полисахариды, выполняющие энергетическую функцию;
Структурные полисахариды, обеспечивающие клеткам и органам механическую прочность;
Полисахариды, входящие в состав межклеточного матрикса, принимают участие в образовании тканей, пролиферации и дифференцировке клеток. Сильно гидратированы.
БИОЛОГИЧЕСКАЯ РОЛЬ УГЛЕВОДОВ:
1. ЭНЕРГЕТИЧЕСКАЯ. При окислении1 г углеводов до конечных продуктов (СО2 и Н2О) выделяется 4,1-ккал энергии. На долю углеводов приходится около 60-70 % всей суточной калорийности пищи. Суточная потребность в углеводах для взрослого человека в среднем массой 60-70 кг составляет около 400-500 г.
2. Анаболическая. Являются источниками субстратов для синтеза жирных кислот и аминокислот.
3. СТРУКТУРНАЯ или пластическая. Углеводы используется как строительный материал для образования структурных компонентов клеток (гликолипиды, гликопротеины, гетерополисахариды межклеточного вещества). Входят в состав нуклеиновых кислот.
4. РЕЗЕРВНАЯ. Углеводы откладываются в клетках в виде резервного полисахарида гликогена.
5. ЗАЩИТНАЯ. Гликопротеины принимают участие в образовании антител. Гиалуроновая кислота, входя в состав соединительной ткани, препятствует проникновению чужеродных веществ. Гетерополисахариды участвуют в образовании вязких секретов покрывающей слизистые оболочки дыхательных путей, мочевыводящих путей, пищеварительного тракта, предохраняя их от повреждений.
6. Обезвреживающая. УДФ-глюкуроновая кислота в печени связывает многие токсические соединения, придавая им большую гидрофильность и способность растворяться в желчи.
5. РЕГУЛЯТОРНАЯ. Некоторые гормоны гипофиза, щитовидной железы являются гликопротеинами. Фукозилированные протеины и пептиды свидетельствуют о канцерогенезе, малигнизации и метастазировании опухолей.
6. Участвуют в процессах узнавания клеток. Важная роль при этом отводятся сиаловым кислотам и нейраминовой кислоте. Гетерополисахариды входя в состав оболочек эритроцитов, определяют группы крови. Входят в состав рецепторов гормонов и нейромедиаторов.
7. Участвуют в процессах свёртывания крови, входят в состав фибриногена и протромбина. Препятствуют свёртыванию крови, входят в состав гепарина.
ПРЕВРАЩЕНИЕ УГЛЕВОДОВ В ПИЩЕВАРИТЕЛЬНОМ ТРАКТЕ
Основными углеводами пищи для организма человека являются: крахмал, гликоген, лактоза, в незначительной степени моносахариды – глюкоза, фруктоза. Для современного человека значимое место стала занимать сахароза.
Переваривание углеводов в ротовой полости. Поступивший с пищей крахмал (гликоген) в ротовой полости подвергается гидролизу под действием альфа-амилазы слюны, которая относится к эндоамилазам. Она расщепляет альфа (1,4)-гликозидные связи в структуре крахмала. рН оптимум для альфа-амилазы слюны находится в слабощелочной среде (рН = 7-8). Так как пища в ротовой полости находится недолго, то крахмал метаболизируется лишь частично. Его гидролиз завершается образованием амилодекстринов. Известны следующие виды декстринов:
амило-декстрины, эритродекстрины, ахродекстрины и. Амилодекстрины (молекулярная масса их около 10000, количество глюкозных единиц около 60);
эритродекстрины - молекулярная масса около 7000, количество глюкозных единиц около 40;
ахродекстрины - молекулярная масса их около 3700, количество глюкозных единиц около 20;
мальтодекстрины – олигосахариды, содержащие от 2 до 20 остатков глюкозы. (malt – солод, англ.)
Далее пища поступает в желудок. Слизистой оболочкой желудка гликозидазы не вырабатываются. В желудке среда резко кислая (рН=1,5-2,5), поэтому действие альфа-амилазы слюны внутри пищевого комка прекращается. Однако в более глубоких слоях действие фермента продолжается, и крахмал успевает пройти следующую стадию гидролиза, с образованием эритродекстринов.
Переваривание углеводов в кишечнике. Основным местом переваривания крахмала служит тонкий отдел кишечника. Здесь проходит наиболее важная фаза гидролиза крахмала. В переваривании крахмала принимает участие ферменты, вырабатываемые в поджелудочной железе (альфа-амилаза, амило-1,6-гликозидаза и олиго-1,6-гликозидаза ).
Выделяющийся панкреатический сок содержит бикарбонаты, которые принимают участие в нейтрализации кислого желудочного содержимого, создаётся слабощелочная среда (рН=8-9) - оптимальная для гликозидаз. Образующиеся катионы (Na+, K+) принимают участие в активации ферментов.
Три панкреатических фермента завершают гидролитический разрыв внутренних гликозидных связей в структуре крахмала. Эритродекстрины переходят в ахродекстрины.
Альфа-амилаза завершает разрыв внутренних альфа-(1,4)-гликозидных связей, амило-1,6-гликозидаза гидролитически расщепляет внутренние альфа-1,6-гликозидные связи в точках ветвления, а олиго-1,6-гликозидаза является терминальной в этом процессе.
Таким образом, три панкреатических фермента завершают гидролиз крахмала в кишечнике с образованием мальтоз (дисахарид, состоящий из двух остатков глюкозы, соединённых α-1,4-гликозидной связью) и изомальтоз (дисахарид, состоящий из двух остатков глюкозы, соединённых α-1,6-гликозидной связью) и триозосахаридов.
В щёточной каёмке тонкого кишечника на наружной поверхности цитоплазматической мембраны энтероцитов расположены ферментные комплексы, расщепляющие гликозидные связи в дисахаридах (дисахаридазы). У человека известно о четырёх ферментно-субстратных комплекса:
сахаразо-изомальтазный комплекс. Прикреплён к мембране микроворсинок с помощью гидрофобного домена. Гидролизует сахарозу и изомальтозу, расщепляя α-1,2- и α-1,6-гликозидные связи, мальтозу и мальтотриозу – гидролизуя α-1,4-гликозидные связи. На его долю приходится 80% всей мальтазной активности кишечника. Сахаразная субъединица – единственный фермент в кишечнике, гидролизующий сахарозу, а изомальтазная субъединица с большей скоростью гидролизует связи в изомальтозе, чем в мальтозе и мальтотриозе. Активность комплекса высокая в тощей кишке, но снижается в проксимальной и дистальных частях кишечника.

Сахаразо-изомальтазный комплекс. 1 – сахараза, 2 - изомальтаза, 3 – связывающий домен, 4 – трансмембранный домен, 5 – цитоплазматический домен.

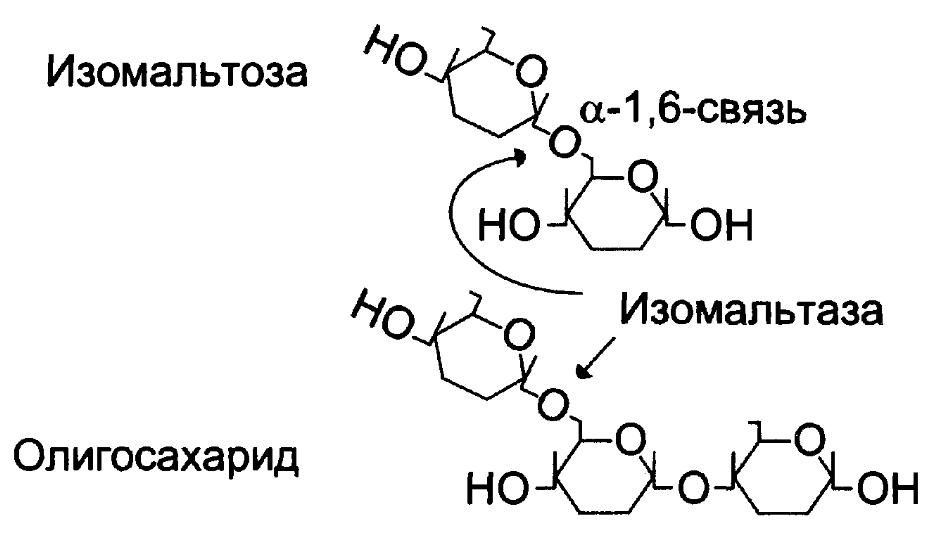
Гликоамилазный комплекс – катализирует гидролиз α-1,4-гликозидные связи между глюкозными остатками в олигосахарах, действуя с восстанавливающего конца – экзогликозидаза. Может действовать как мальтаза. Активность наибольшая в нижних отделах тонкого кишечника.
β-гликозилазный комплекс (лактаза) – расщепляет β-1,4-гликозидную связь в молочном сахаре между галактозой и глюкозой. Является гликопротеином. Неравномерно распределён к кишечнике. Наиболее активен у плода в последние сроки беременности и до 5-7 летнего возраста. С возрастом – активность снижается (у взрослых составляет до 10% от детского возраста) у некоторых исчезает вовсе.

Трегалаза – гидролизует связь между мономерами трегалозы – дисахариде грибов состоящей из двух остатков глюкозы, связанных гликозидной связью между первыми аномерными атомами углерода.

Транспорт глюкозы.
Гидролиз соответствующих дисахаридов пищи заканчивается образованием мономеров - глюкозы, галактозы, фруктозы, которые всасываются эпителиальными клетками тощей и подвздошной кишок с помощью специальных механизмов транспорта через мембраны этих клеток:
путём облегчённой диффузии; Облегчённую диффузию обеспечивают белки-переносчики семейства ГЛУТ (GLUT или SLC2A). Они имеют 12 чрезмембранных спиральных доменов. Считается, что 3,5,7 и 11 трансмембранные сегменты образуют канал для глюкозы.
Глюкозные транспортёры (GLUT) обнаружены во всех тканях. Существует несколько разновидностей ГЛЮТ, они пронумерованы в соответствии с порядком их обнаружения. В настоящее время описанны 12 типов GLUT имеют сходные первичную структуру и доменную организацию.
ГЛЮТ-1 - широко распространён в клетках тканей плода, в эритроцитах, эндотелиоцитах гематоэнцефалического барьера. количество GLUT-1 повышается при низком уровне глюкозы.
ГЛЮТ-2 обнаружен в клетках канальцев почек, энтероцитах тонкого кишечника, клетках печени и β-клетках поджелудочной железы (играет роль сенсора глюкозы). Обеспечивает двунапраленное перемещение – при гликолизе – внутрь, при глюконеогенезе – наружу. Из энтероцитов в кровь ГЛУТ-2 переносит глюкозу, фруктозу и галактозу;
ГЛЮТ-3 – синтезируется в основном в нейронах (основная форма) и плаценты. Обладает высоким сродством к глюкозе;
ГЛЮТ-4 - главный переносчик глюкозы в клетки мышц (в т.ч. в кардиомиоциты) и жировой ткани. Контролируется инсулином; который стимулирует транслокацию специфичных GLUT-4 содержащих пузырьков из внутриклеточного пространства к плазматической мембране, что способствует немедленному увеличению транспорта глюкозы в 10-20 раз.
ГЛЮТ-5 - транспортирует только фруктозу. Встречается, главным образом, в клетках тонкого кишечника, яичках и почках.
ГЛЮТ-7 – при распаде гликогена в лизосомах (гликогенолизе) транспортирует глюкозу из эндоплазматической сети.
Все типы ГЛЮТ могут находиться как в плазматической мембране, так и в цитозольных везикулах. ГЛЮТ-4 (и в меньшей мере ГЛЮТ-1) почти полностью находятся в цитоплазме клеток. Влияние инсулина на такие клетки приводит к перемещению везикул, содержащих ГЛЮТ, к плазматической мембране, слиянию с ней и встраиванию транспортёров в мембрану. После чего возможен облегчённый транспорт глюкозы в эти клетки. После снижения концентрации инсулина в крови транспортёры глюкозы снова перемещаются в цитоплазму, и поступление глюкозы в клетку прекращается (рис. 7-19). Известны различные нарушения в работе транспортёров глюкозы. Наследственный дефект этих белков может лежать в основе инсулинонезависимого сахарного диабета. В то же время причиной нарушения работы транспортёра глюкозы может быть не только дефект самого белка. Нарушения функции ГЛЮТ-4 возможны на следующих этапах:
передача сигнала инсулина о перемещении этого транспортёра к мембране;
перемещение транспортёра в цитоплазме;
включение в состав мембраны;
отшнуровывание от мембраны и т.д.
После всасывания моносахариды покидают клетки слизистой оболочки кишечника через мембрану, обращённую к кровеносному капилляру, с помощью облегчённой диффузии.
Потребление глюкозы клетками из кровотока происходит так же с помощью облегчённой диффузии. Исключение составляют клетки мышц и жировой ткани, у которых облегчённая диффузия регулируется инсулином.
путём активного транспорта (вторичный активный транспорт). Глюкоза и Na+ проходят через мембраны с люминальной стороны, связываясь с различными участками белка-переносчика (НГЛТ – натрий зависимый транспортёр глюкозы) (sodium-dependent glucose transport (SGLT; SGLT-1, -2,-3). При этом натрий поступает в клетку по градиенту концентрации, и одновременно глюкоза транспортируется против градиента концентрации (вторичный активный транспорт). Т.е чем больше градиент Na+ - тем больше поступление глюкозы в энтероциты и наоборот. Градиент концентрации Na+ - движущая сила активного симпорта, создаётся работой Na+,K+-АТФ-азой. Такой же механизм переноса и для галактозы. Активный транспорт особенно эффективен при низкой концентрации глюкозы. Если же концентрация глюкозы (фруктозы) в кишечнике велика, то она может транспортироваться в клетку путём облегчённой диффузии. Всасывание глюкозы и галактозы гораздо выше, чем других моносахаридов.
Продукты полного гидролиза - моносахариды - всасываются в кровь и на этом завершается начальный этап обмена углеводов - пищеварение.
С пищей в организм человека поступает клетчатка, которая в пищеварительном тракте не переваривается, поскольку отсутствуют бета -гликозидазы.
Однако биологическая роль клетчатки велика: она формирует пищевой комок, продвигаясь по желудочно-кишечному тракту она:
нормализует кишечную микрофлору (место обитания ~ 3 кг микроорганизмов);
раздражает слизистые оболочки, усиливая сокоотделение;
клетчатка усиливает перистальтику кишечника;
достигая отделов толстого кишечника клетчатка под действием ферментов условно-патогенной микрофлоры подвергается брожению с образованием глюкозы, лактозы и газообразных веществ.

Метаболизм глюкозы
1,
2,
3,
Анаэробный гликолиз
В зависимости от функционального состояния организма, клетки органов и тканей могут находиться как в условиях достаточного снабжения кислородом, так и испытывать его недостаток, то есть находится в условиях гипоксии. Если катаболизму подвергается глюкоза, то процесс называется ГЛИКОЛИЗОМ, если распадается глюкозный остаток гликогена –ГЛИКОГЕНОЛИЗОМ. В связи с этим катаболизм углеводов может рассматриваться с двух позиций:
1.В АНАЭРОБНЫХ УСЛОВИЯХ
2.В АЭРОБНЫХ УСЛОВИЯХ.
АНАЭРОБНЫЙ ГЛИКОЛИЗ протекает в цитоплазме клеток. Окисление глюкозы всегда завершается образованием конечного продукта этого процесса - молочной кислоты.
Окисление глюкозы в тканях начинается с образования глюкозо-6-фосфата. Дальнейшее окисление углеводов в тканях, как в ана-, так и в аэробных условиях полностью совпадает до стадии образования пирувата.
Процесс анаэробного гликолиза сложный и многоступенчатый. Условно его можно разделить на 2 стадии:
-первая стадия заканчивается образованием из гексозы двух триоз: -диоксиацетонфосфата и глицеральдегид-3-фосфата.
-Вторая стадия называется стадией гликолитической оксидоредукции. Эта стадия катаболизма наиболее важная, поскольку она сопряжена с образованием АТФ, за счёт реакций субстратного фосфорилирования, окислением глицральдегид -3-фосфата, восстановлением пирувата до лактата.

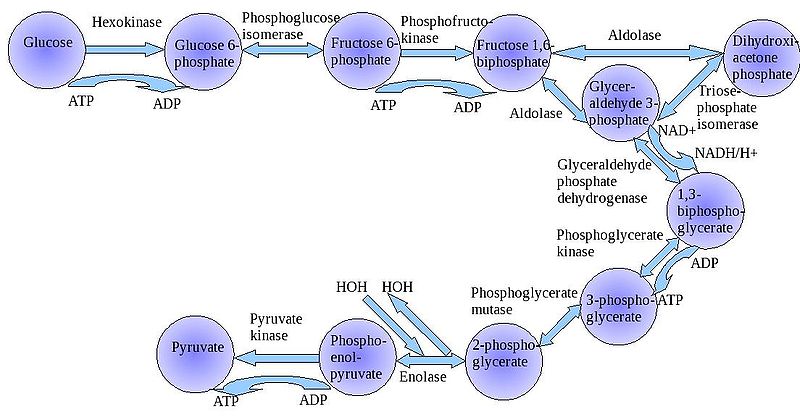
В процессе окисления глюкозы было израсходовано 2 молекулы АТФ (гексокиназная и фосфофруктокиназная реакции). С этапа образования триоз идёт одновременное их окисление. В результате этих реакций образуется энергия в виде АТФ за счёт реакций субстратного фосфорилирования (глицераткиназная и пируваткиназная реакции).
На этапе гликолитической оксидоредукции идёт окисление глицеральдегид-3-фосфата в присутствии НЗРО4 и НАД-зависимой дегидрогеназы, которая при этом восстанавливается до НАДН2. Митохондрии в анаэробных условиях блокированы, поэтому выделенные в результате окисления молекулы НАДН2 находится в среде до тех пор, пока не образуется субстрат, способный принять их. Пируват, принимая НАДН2, восстанавливается до лактата, завершая тем самым внутренний-окислительно-восстановительный этап гликолиза. НАД- окисленный выделяется и может вновь участвовать в окислительном процессе, выполняя роль переносчиков водорода.
3 реакции гликолиза являются необратимыми:
1. гексокиназная.
2. фосфофруктокиназная.
3. пируваткиназная.
Энергетический эффект окисления 1 молеклы глюкозы составляет 2 АТФ.
Биологическая роль анаэробного гликолиза - энергетическая.
Анаэробный гликолиз является единственным процессом, продуцирующим энергию в форме АТФ в клетке в бескислородных условиях. В эритроцитах гликолиз является единственным процессом, продуцирующим АТФ и поддерживающим биоэнергетику, для сохранения их функции и целостности.

Аэробный гликолиз (гексозодифосфатный путь)
Это классический путь аэробного катаболизма углеводов в тканях протекает в цитоплазме до стадии образования пирувата и завершается в митохондриях с образование конечных продуктов АТФ и Н2О и выделением CO2.
Когда в клетки начинает поступать кислород - происходит подавление анаэробного гликолиза. При этом снижается потребление глюкозы, блокируется образование лактата. Эффект торможения анаэробного гликолиза дыханием получил название эффекта Пастера. Окисление углеводов до стадии образования пирувата происходит в цитоплазме клеток. Затем пируват поступает в митохондрии, где в матриксе подвергается дальнейшему окислению. В результате реакции окислительного декарбоксилирования образуется ацетил-КоА который, в дальнейшем окисляется с участием ферментов цикла Кребса и сопряженных с ним ферментов цепи переноса электронов (ЦПЭ).
Происходит образование конечных продуктов (СО2 и Н2О), выделяется энергия в форме АТФ.
Н2О образуется на этапе превращения:
1. ГЛИЦЕРАЛЬДЕГИД-3-ФОСФАТА
2. 2-ФОСФОГЛИЦЕРИНОВОЙ КИСЛОТЫ
3. ПИРУВАТА
4. Альфа- КЕТОГЛУТАРОВОЙ КИСЛОТЫ
5. СУКЦИНАТА
4. ИЗОЦИТРАТА
7. МАЛАТА
СО2 образуется на этапе превращения:
1. ПИРУВATА
2. ОКСАЛОСУКЦИНАТА
3. Альфа - КЕТОГЛУТАРОВОЙ КИСЛОТЫ.
АТФ образуется:
А. За счёт реакций СУБСТРАТНОГО ФОСФОРИЛИРОВАНИЯ на этапе превращения:
1. 1,3-ДИФОСФОГЛИЦЕРИНОВОЙ К-ТЫ
2. 2-ФОСФОЕНОЛПИРУВАТА
3. СУКЦИНИЛА-КОА
В. За счёт реакций ОКИСЛИТЕЛЬНОГОФОСФОРИЛИРОВАНИЯ на этапе превращения:
1. ГЛИЦЕРАЛЬДЕГИД-3-ФОСФАТА
2. ПИРУВАТА
3. ИЗОЦИТРАТА
4. альфа – КЕТОГЛУТАРОВОЙ КИСЛОТЫ
5. СУКЦИНАТА
6. МАЛАТА.
Энергетический эффект окисления глюкозы в аэробных условиях составляет 38 АТФ, глюкозного остатка гликогена 39 АТФ.