

ОБМЕН УГЛЕВОДОВ
Гликоген, строение , биороль ,синтез, мобилизация
Строение. Гликоген - главная форма запасания углеводов у животных, в растениях эту функцию выполняет крахмал. Наибольшие запасы гликогена имеются в печени (до 6% массы печени) и в мышцах (до 1% мышечной массы). Продолжительность использования гликогена зависит от степени физической активности. В покое запас гликогена расходуется примерно за 18-20 часов, при выполнении физической работы за 2-4 часа. Практически не имеют гликогена клетки нервной ткани. Они целиком зависят от циркулирующей в крови глюкозы.
Гликоген представляет собой разветвленный гомополисахарид, построенный из сотен тысяч остатков Д-глюкозы, соединенных в полисахаридные цепи -1,4-гликозидными связями, а в местах ветвления -1,6-гликозидными связями. Линейные участки между точками содержат 12-18 остатков глюкозы.
Молекулярная масса гликогена колеблется от 106-109Д
Биороль. Функции гликогена печени и других органов существенно отличаются. Гликоген мышц является легкодоступным источником глюкозы, которая используется в самой мышце для покрытия энергетических нужд. Гликоген печени используется главным образом для поддержания физиологической концентрации глюкозы в крови в промежутках между приемами пищи.
Биосинтез. Для синтеза гликогена используется глюкоза крови при условии если ее концентрация выше физиологической нормы.. Синтез гликогена осуществляется в два этапа. На первом этапе происходит образование активной формы глюкозы, второй этап заключается в наращивании цепей затравочного гликогена. В ходе реакций первого этапа глюкоза вначале фосфорилируется по 6 положению под действием гексокиназы в мышцах или глюкокиназы в печени. Донором фосфатной группы является АТФ.
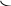
 АТФ АДФ
АТФ АДФ
 Глюкоза
Глюкозо-6-фосфат
Глюкоза
Глюкозо-6-фосфат
В следующей реакции под действием фосфоглюкомутазы происходит внутримолекулярный перенос фосфата из 6 положения в первое
 глюкозо-6-фосфат
глюкозо-1-фосфат
глюкозо-6-фосфат
глюкозо-1-фосфат
Далее глюкозо-1-фосфат под действием глюкозо-1-фосфатуридинтрансферазы реагирует с уридинтрифосфатом, в результате образуется активный нуклеотид уридиндифосфатглюкоза (УДФ-глюкоза)
 УТФ +
глюкозо-1-фосфат УДФ-глюкоза
+ РР
УТФ +
глюкозо-1-фосфат УДФ-глюкоза
+ РР
На этой реакции заканчивается первый этап образования активной формы глюкозы. На втором этапе под действием фермента гликогенсинтазы образуется гликозидная связь между атомом С1 активированной глюкозы и атомом С4 остатка глюкозы в гликогене.
УДФ-глюкоза + (С6)n УДФ + (С6)n+1
В етви
гликогенового дерева удлиняются путем
последовательного образования 1 4
гликозидных связей. После того как длина
линейного участка достигнет как минимум
11 остатков глюкозы ветвящий или
бранчинг-фермент переносит фрагмент
длиной в 6 остатков на соседнюю цепь,
присоединяя его
етви
гликогенового дерева удлиняются путем
последовательного образования 1 4
гликозидных связей. После того как длина
линейного участка достигнет как минимум
11 остатков глюкозы ветвящий или
бранчинг-фермент переносит фрагмент
длиной в 6 остатков на соседнюю цепь,
присоединяя его
1 6 гликозидной связью, таким образом
образуется точка ветвления. В дальнейшем
ветви растут путем последовательного
присоединения глюкозных остатков. По
мере ветвления скорость синтеза гликогена
возрастает. Путь Кристмас.и Лелуара
6 гликозидной связью, таким образом
образуется точка ветвления. В дальнейшем
ветви растут путем последовательного
присоединения глюкозных остатков. По
мере ветвления скорость синтеза гликогена
возрастает. Путь Кристмас.и Лелуара
Распад гликогена происходит под действием фермента гликогенфосфорилазы, катализирующего фосфоролитическое расщепление связей 14 , продуктом является глюкозо-1-фосфат.
(С6)n + Р (С6)n-1 + глюкозо-1-фосфат
Остатки глюкозы отщепляются до тех пор, пока на ветви не останется 4 глюкозных остатка. Специальная глюкантрансфераза переносит трисахаридный фрагмент с одной цепи на другую, открывая пункт ветвления. Гидролитическое расщепление 16 связи осуществляет деветвящий или дебрачинг фермент с образованием свободной глюкозы. Образовавшаяся в результате фосфоролиза глюкозо-1-фосфат изомеризуется в глюкозо-6-фосфат, который в мышцах включается в гликолитическую цепь реакций.
Глюкозо-1-фосфат глюкозо-6-фосфат
В печени и почках имеется фермент глюкозо-6-фосфатаза, который отщепляет фосфат от глюкозо-6-фосфата с образованием свободной глюкозы, которая диффундирует из клеток в кровь
глюкозо-6-фосфат глюкоза + фосфат. Амилолиз заключается в отщеплении остатков глюкозы под действием амилазы.
АНАЭРОБНЫЕ ПУТИ ПРЕВРАЩЕНИЯ УГЛЕВОДОВ. ГЛИКОЛИЗ
СПИРТОВОЕ БРОЖЕНИЕ.
Гликолиз или молочнокислое брожение - анаэробный распад глюкозы до лактата ради энергии. Гликолиз является наиболее древним биохимическим процессом, так как первичная атмосфера земли не содержала кислорода.
Совершается в тканях животных в условиях кислородной недостаточности. В эритроцитах, где отсутствуют митохондрии, гликолиз является единственным процессом обеспечивающим образование АТФ. Высокая интенсивность гликолитического распада углеводов отмечается в эмбриональной и опухолевой ткани. Процесс является уникальным так как является главным путем метаболизма не только глюкозы , но и фруктозы и галактозы.
Гликолиз представляет собой линейную последовательность 11 реакций, протекающих в цитозоле клеток и является примером функциональной формы организации полиферментных систем. Гликолиз состоит из двух этапов: первый этап является подготовительным, в ходе которого глюкоза дважды фосфорилируется и подвергается дихотомии с образованием двух молекул глицеральдегид-3-фосфата.
Второй этап это гликолитическая оксидоредукция. В ходе реакций этого этапа происходят реакции, приводящие к накоплению в метаболитах энергии в виде макроэргических связей, за счет разрыва которых происходит две реакции субстратного фосфорилирования и образуется АТФ.
Суммарное уравнение гликолиза следующее
глюкоза + 2АДФ + 2Р 2 лактат + 2АТФ + Н2О
Гликолитический путь превращения глюкозы начинается с ее фосфорилирования по 6 положению под действием гексокиназы. Донором фосфатной группы выступает АТФ. Реакция характеризуется значительными потерями энергии в виде теплоты, поэтому она является необратимой. Продуктом реакции является глюкозо-6-фосфат , неспособный покинуть клетку в силу наличия отрицательного заряда. Глюкозо-6-фосфат занимает важное положение в области стыковки ряда метаболических путей ( гликолиз, гликогенолиз, глюконеогенез, пентозофосфатный путь. В ходе гликолиза он превращается в во фруктозо-6-фосфат при участии фосфогексозоизомеразы , т.е. альдоза в реакции изомеризации превращается в кетозу.
Далее следует еще одно фосфорилирование, которое катализируется фосфофруктокиназой с образованием фруктозо-1,6-дифосфата
 Фруктозо-1-фосфат
+ АТФ фруктозо-1,6-дифосфат
+ АДФ
Фруктозо-1-фосфат
+ АТФ фруктозо-1,6-дифосфат
+ АДФ
Донором фосфатной группы является АТФ. Реакция является необратимой. Фосфофруктокиназа является ключевым ферментом гликолиза, именно он определяет скорость всех остальных реакций. Его активность увеличивается в присутствии АМФ, фруктозо-6-фосфата, а подавляется АТФ и цитратом.
Заканчивается первый этап альдольным расщеплением фруктозо-1,6-дифосфата под действием альдолазы на два триозофосфата : глицеральдегид-3-фосфат и дигидроксиацетонфосфат ,т.е. исходная гексоза располовинивается на две равные части.
 Фруктозо-1,6-фосфат
глицеральдегид-3-фосфат +
дигидроксиацетонфосфат.
Фруктозо-1,6-фосфат
глицеральдегид-3-фосфат +
дигидроксиацетонфосфат.
Глицеральдегид-3-фосфат и дигидроксиацетонфосфат превращаются друг в друга при участии триозофосфатизомеразы
 Дигидоксиацетонфосфат
глицеральдегид-3-фосфат
Дигидоксиацетонфосфат
глицеральдегид-3-фосфат
В дальнейших реакциях гликолиза может участвовать только глицеральдегид-3-фосфат.
Окислительно-восстановительный этап начинается с окисления глицеральдегид-3-фосфата с образованием 1,3-дифосфоглицерата. Дигидроксиацетонфосфат также окисляется в 1,3-дифосфоглицерат, проходя через стадию образования глицеральдегид-3-фосфата.
 Глицеральдегид-3-фосфат
+ НАД + Р 1,3-дифосфоглицерат
+ НАДН2
Глицеральдегид-3-фосфат
+ НАД + Р 1,3-дифосфоглицерат
+ НАДН2
Реакция катализируется глицеральдегидфосфатдегидрогеназой. Акцептором атомов водорода в этой реакции является НАД (производное витамина РР). В результате фосфорилирования у первого углеродного атома образуется макроэргическая связь. Высокоэнергетический фосфат переходит в следующей реакции в состав АТФ при участии фермента фосфоглицераткиназы при этом образуется 3-фосфоглицерат. Эта реакция является одной из реакций субстратного фосфорилирования

 1,3-дифосфоглицерат
+ АДФ 3-фосфоглицерат + АТФ
1,3-дифосфоглицерат
+ АДФ 3-фосфоглицерат + АТФ
Поскольку на каждую молекулу глюкозы, участвующую в гликолизе, образуется две молекулы триозы, то на данной стадии образуется две молекулы АТФ на молекулу глюкозы. В качестве разобщителя субстратного фосфорилирования на данном этапе выступает арсенат.
3-фосфоглицерат в ходе внутримолекулярного переноса фосфата превращается в 2-фосфоглицерат под действием фермента фосфоглицератмутазы.
На следующей стадии, катализируемой енолазой, происходит отщепление молекулы воды и перераспредление энергии внутри молекулы, при этом фосфат в положении 2 переходит в высокоэнергетическое состояние. Продуктом реакции является фосфоенолпируват
2-фосфоглицерат фосфоенолпируват + Н20
Высокоэнергетический фосфат фосфоенолпирувата переносится на АДФ ферментом пируваткиназой, т.е. происходит субстратное фосфорилирование. На этой стадии образуются еще две молекулы АТФ на молекулу глюкозы. Продуктом реакции является пируват. Реакция является необратимой
Фосфоенолпируват + АДФ пируват + АТФ
На заключительном этапе происходит восстановление пирувата в лактат под действием лактатдегидрогеназы. В этой реакции используется восстановленный НАДН, образовавшийся в шестой реакции гликолиза (глицеральдегиддегидрогеназной реакции), который окисляется до НАД. Эта реакция обеспечивает регенерацию окисленного НАД, необходимого для осуществления шестой реакции гликолиза. Благодаря наличию двух окислительно-восстановительных реакций исходный субстрат – глюкоза и конечный продукт гликолиза- лактат имеют одинаковую степень окисленности.
Пируват + НАДН2 лактат + НАД
Энергетическая ценность гликолиза составляет 2 АТФ; на подготовительном этапе расходуется две молекулы АТФ в реакциях фосфорилирования глюкозы и фруктоз-6-фосфата, а на этапе гликолитической оксидоредукции происходит образование 4 АТФ в двух реакциях субстратного фосфорилирования.
Спиртовое брожение отличается от гликолиза двумя заключительными реакциями. Пируват в этом случае декарбоксилируется в образованием ацетальдегида. Реакция катализируется пирувадекарбоксилазой и требует присутсивя ТПФ (производное витамина В1). В следующей реакции происходит восстановление ацетальдегида под действием алкогольдегидрогеназы, использующей восстановленный НАД, с образованием этанола. Реакции спиртового брожения катализируются ферментным комплексом из 12 ферментов микрорганизмов, пивных дрожжей, который называется зимазой.
Гликогенолиз отличается двуми первыми реакциями, приводящими к образованию глюкозо-6-фосфата. С этими рекциями вы уже познакомились при рассмотрении мобилизации гликогена.
Значение гликолиза сводится к возможности получения АТФ в анаэробных условиях, т.е. это аврийный путь, включающийся в условиях гипоксии. В энергетическом отношении путь крайне не эффективен, но в то же время это единственный путь дающий энергию в анаэробных условиях.
Пластическое значение гликолитической цепи заключается в том, что некоторые метаболиты могут использоваться в реакциях синтеза. Например, дигидроксиацетонфосфат – на синтез липидов, пируват на образование аланина.
Гликолитическая цепь до стадии образования пирувата представляет собой первый этап дихотомического распада углеводов.
Аэробный дихотомический распад углеводов
Представляет собой сложный, многоступенчатый, необратимый процесс аэробного расщепления углеводов до конечных продуктов СО2 и воды. Условно аэробный дихотомический распад можно разделить на три этапа.
Первый этап представляет собой 10 реакций гликолиза до стадии пирувата. Расщепление глюкозы на этом этапе происходит через промежуточные продукты гликолиза, используются ферменты гликолиза. Отличительной ососбенностью является механизм окисления НАДН, образовавшегося в глицеральдегиддегидрогеназной реакции. В гликолизе он окисляется в лактатдегидрогеназной реакции за счет восстановления пирувата до лактата. В аэробных условиях окисление НАДН обеспечивается с помощью челночных механизмов транспорта водорода в митохондрии. Существует несколько таких систем собирания водорода и переноса его в виде метаболитов через мемебраны митохондрий. Благодаря челночным механизмам водород с цитоплазматического НАД переносится на митохондриальный НАД, окисленный НАД может вновь включиться в реакцию окисления глицеральдегид-3-фосфата. При этом восстановление пирувата в лактат не происходит, так как это возможно только в присутствии восстановленного НАД. Поэтому в аэробных условиях пируват подвергается непрямому окислительному декарбоксилированию под действием ферментного комплекса приуватдегидрогеназы. Это превращение выделяется как
Второй этап аэробного распада углеводов. Продуктом реакции является ацетил-КоА. Реакция требует присутствия таких коферментом как ТПФ (производное вит В1, отвечает за декарбоксилирование пирувата), ФАД ( производное вит В2, является промежуточным акцептором водорода ), НАД ( производное вит РР, является конечным акцептором водорода), КоА ( производное пантотеновой кислоты, отвечает за образование активной формы уксусной кислоты), липоевая кислота необхо димая для фиксации субстрата реакции. Образовавшийся ацетил-КоА включается в цикл трикарбоновых кислот, который в данном случае служит
Третьим этапом аэробного распада глюкозы.
Энергетическая эффективность аэробного дихотомического распада.
1 этап
Глицеральдегидфосфатдегидрогеназная реакция 2 восст НАД 6 АТФ
Глицераткиназная реакция 2 реакци субст фосф 2 АТФ
Пируваткиназная реакция 2 реакции субст фосф 2АТФ
Всего образовалось 10АТФ
Израсходовано на фосфорилирование глюкозы 2 АТФ
2 этап
пируватдегидрогеназная реакция 2 восст НАД 6 АТФ
3 этап
изоцитратдегидрогеназная реакция 2 восст НАД 6 АТФ
2-кетоглутаратдегидрогеназная реакция 2 восст НАД 6 АТФ
сукцинил-КоА-синтазная реакция 2 реакции субст фосф 2 АТФ
сукцинатдегидрогеназная реакция 2 восст ФАД 4 АТФ
малатдегидрогеназная реакция 2 восст НАД 6 АТФ
всего образовалось 24 АТФ