

5.3.1.Парадокс возможности существования многоклеточных организмов
Огромный размер генома многоклеточных организмов с генетической точки зрения должен создавать для их существования многочисленные и, на первый взгляд, трудноразрешимые препятствия. Проблемы начинаются уже при редупликации гигантских молекул геномной ДНК с помощью ферментных систем, точность функционирования которых не является абсолютной. Кроме того, репликация происходит в присутствии мутагенов экзогенного и эндогенного происхождения. Принято считать, что частота спонтанных мутаций в геноме соматических клеток млекопитающих, растущих в культуре, варьирует от локуса к локусу одного и того же генома и, по разным оценкам, достигает значений от 10-8 до 10-12 на нуклеотид за клеточную генерацию. Точное определение частоты спонтанных мутаций в геномной ДНК высших эукариот in vivo является сложной задачей. По ряду экспериментальных и косвенных данных, в частности, исходя из частоты встречаемости некоторых наследственных заболеваний в популяциях человека (например серповидно-клеточной анемии, вызываемой заменой единственного нуклеотида в геномной ДНК), полагают, что эти показатели, по крайней мере, не ниже значений, полученных in vitro.
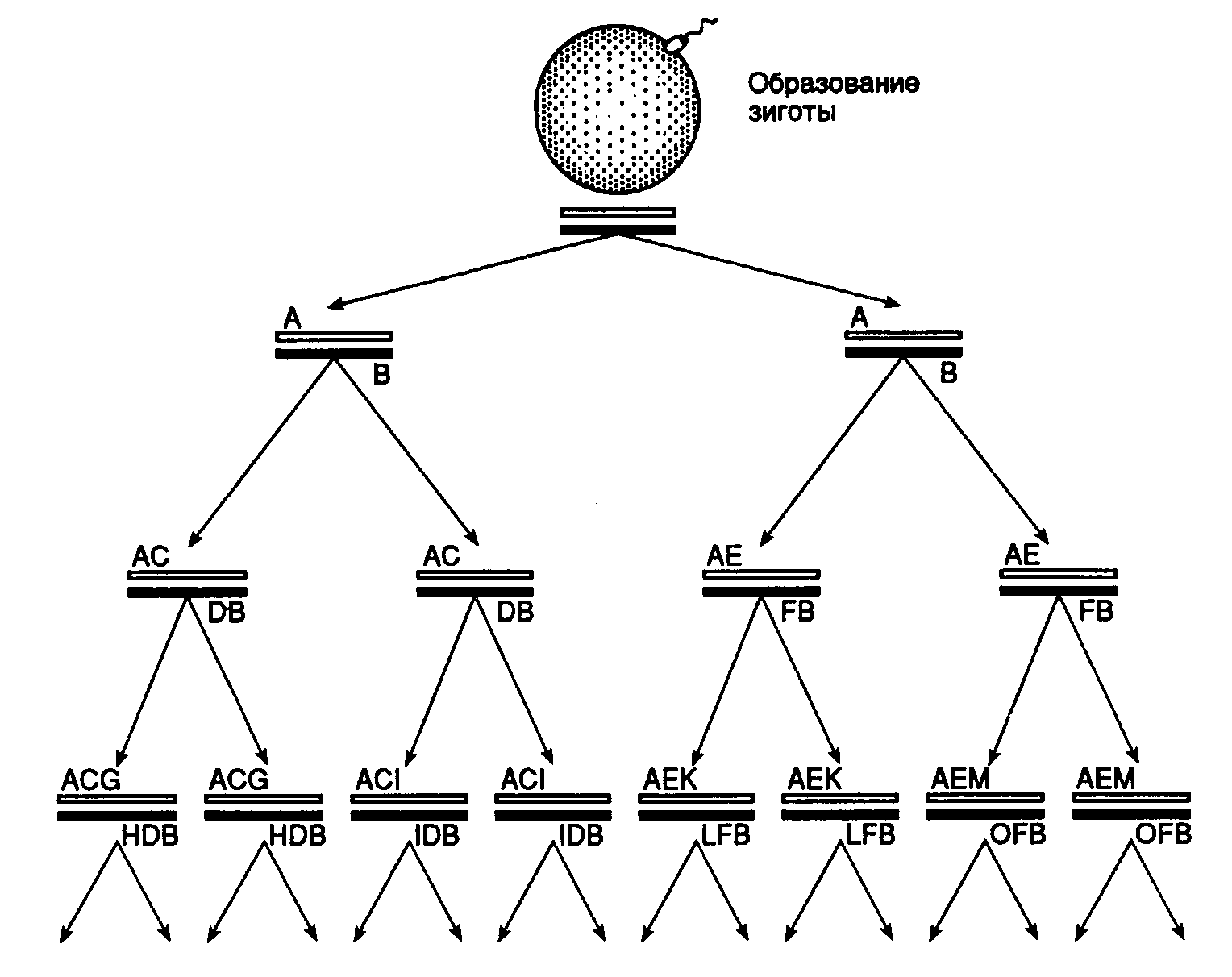
Рис. I.62. Происхождение генетического груза в геноме соматических клеток многоклеточных организмов
Развитие многоклеточного организма начинается с дробления зиготы, образующейся в процессе оплодотворения яйцеклетки и содержащей диплоидный набор хромосом (две параллельные линии вверху рисунка). А–М – наборы мутаций
Принимая, что суммарная ДНК гаплоидного генома человека насчитывает 3·109 п.о., а частота спонтанных мутаций в среднем составляет 10-8 на нуклеотид за генерацию, можно предположить, что, начиная с первого деления оплодотворенной яйцеклетки в процессе онтогенетического развития организма человека, каждое следующее деление должно сопровождаться появлением в их геномной ДНК, по крайней мере, 30 независимых мутаций (рис. I.62). Организм человека состоит из 1015 клеток. Для образования стольких клеток из оплодотворенной яйцеклетки требуется 50 клеточных генераций. Следовательно, гаплоидный геном каждой из соматических клеток человека 50-й генерации должен содержать в разных частях, по крайней мере, 1500 мутаций. Если предположить далее, что набор из 30 мутаций возникает и закрепляется в каждом последующем клеточном делении независимо от мутаций, полученных во время предыдущих делений клеток, то получается, что любая дочерняя соматическая клетка наследует от клетки-предшественницы все имеющиеся в ее геноме мутации и приобретает блок новых 30 мутаций. При этом набор соматических мутаций в потомстве каждой из делящихся клеток одного поколения будет отличаться один от другого, в том числе и в парах аллельных локусов ДНК отдельных клеток, поскольку они редуплицируются независимо. При таком развитии событий мутации, возникающие в геноме соматических клеток каждой последующей генерации, случайным образом сканируют шаг за шагом всю реплицирующуюся геномную ДНК, а результаты сканирования не повторяются в каждом новом поколении клеток. Так, блок из 30 спонтанных мутаций, возникающих после гипотетической 50-й генерации соматических клеток в онтогенезе многоклеточного организма, должен присутствовать в 1015 (250) вариантах.
Проблема становится еще более очевидной, если иметь в виду, что многие клетки организма, например эпителиальные или стволовые клетки крови, пролиферируют на протяжении всей жизни многоклеточного организма, совершая громадное число клеточных делений. По некоторым оценкам, общее число клеточных циклов, в которых участвуют клетки человека на протяжении его жизни, приближается к 1016. Кроме того, соматические мутации возникают в многоклеточном организме не только в активно пролиферирующих, но и покоящихся клетках. Частоты возникновения спонтанных мутаций в пролиферирующих и в покоящихся клетках млекопитающих в ряде случаев различаются лишь незначительно.
Парадоксальность эволюционно сложившейся генетической ситуации заключается в том, что если бы большая часть последовательностей нуклеотидов геномной ДНК заключала в себе жизненно важную генетическую информацию, существование многоклеточных организмов было бы невозможно. Их гибель происходила бы из-за неизбежного накопления в делящихся соматических клетках вредных или летальных мутаций, приводящих к обрыву линий дифференцирующихся в онтогенезе соматических клеток. Это особенно относится к мутациям в жизненно важных генах половых хромосом, которые в соматических клетках находятся в гемизиготном состоянии. Альтернативно основные генетические локусы многоклеточных организмов могут находиться под дополнительной защитой от мутационных изменений.2
На основании приведенных выше аргументов можно сделать вывод о том, что большая часть некодирующих избыточных последовательностей нуклеотидов геномной ДНК эукариот не заключает в себе жизненно важной генетической информации и/или у эукариотических клеток имеются механизмы, обеспечивающие дополнительную защиту геномной ДНК от мутаций. Не соглашаясь с основными положениями концепции "эгоистической" и "паразитической" ДНК, можно предположить, что именно такая организация генома эукариотических организмов, в корне отличающаяся от структуры генома прокариот, имеет отношение к разрешению проблемы генетической парадоксальности существования многоклеточных организмов, которую можно назвать парадоксом М (Metazoa, Metaphyta). Создается впечатление, что именно избыточная ДНК генома эукариот может иметь отношение к повышению его информационной стабильности до уровня, необходимого для реализации многоклеточности в природе.