

2.4.Вектори на основі ретровірусів
Г
Рис. 10 Ретровірус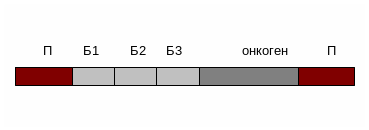
Ретровірусні вектори широко використовуються для доставки генетичного матеріалу в геном людини. Специфічність взаємодії ретровірусів з поверхнею відповідних клітин в момент ураження визначається білком оболонки віріона, який взаємодіє з білками клітини. Це робить можливою зміну клітин-господарів.
Приклади задач
1. Як вектор для клонування була використана плазміда pBR322. Відомо, що донорну ДНК і векторну плазміду попередньо було оброблено рестриктазою PstI з утворенням рекомбінантної плазміди. Які операції треба зробити, щоб ідентифікувати бактеріальні клітини, трансформовані рекомбінантною плазмідою?
Розв’язання.
З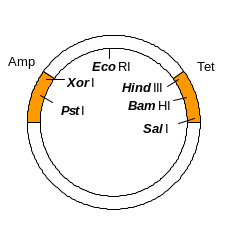 генетичної карти плазміди pBR322
бачимо, що сайт рестрикції PstI
знаходиться в гені Amp,
відповідно рекомбінантні плазміди і
трансформовані ними бактерії втратили
стійкість до ампіциліну. На першому
етапі ідентифікації бактеріальні
клітині висівають на штучне середовище,
що містить тетрациклін. В результаті
виживають тільки ті клітини, які містять
плазміди (інтактні або рекомбінантні).
Другим етапом буде висівання бактеріальних
клітин на середовище з ампіциліном, в
наслідок цього загинуть клітини, що
містять рекомбінантні плазміди, оскільки
вони втратили стійкість до цього
антибіотика. Живими залишаються клітини,
трансформовані нормальною плазмідою
pBR322з
генами стійкості до ампіциліну та
тетрацикліну.
генетичної карти плазміди pBR322
бачимо, що сайт рестрикції PstI
знаходиться в гені Amp,
відповідно рекомбінантні плазміди і
трансформовані ними бактерії втратили
стійкість до ампіциліну. На першому
етапі ідентифікації бактеріальні
клітині висівають на штучне середовище,
що містить тетрациклін. В результаті
виживають тільки ті клітини, які містять
плазміди (інтактні або рекомбінантні).
Другим етапом буде висівання бактеріальних
клітин на середовище з ампіциліном, в
наслідок цього загинуть клітини, що
містять рекомбінантні плазміди, оскільки
вони втратили стійкість до цього
антибіотика. Живими залишаються клітини,
трансформовані нормальною плазмідою
pBR322з
генами стійкості до ампіциліну та
тетрацикліну.
2. Для клонування фрагменту ДНК використали вектор серії EMBL на основі фага λ. Генетична карта вектора має такий вигляд:


 SalGI
BamHI
EcoRI
EcoRI
BamHI
SalGI
SalGI
BamHI
EcoRI
EcoRI
BamHI
SalGI



Для того, щоб не відділяти під час підготовки до роботи „плечі” вектора від центральної частини, вектор обробляють рестриктазою EcoRI, а фрагменти, що утворились – рестриктазою BamHI. Поясніть, для чого це робиться та зазначте які „липкі”кінці має фрагмент чужорідної ДНК, що клонується.
Розв’язання.
Для того, щоб не відділяти під час підготовки до роботи „плечі” вектора від його центральної частини, необхідно, щоб „ липкі” кінці „плеч” вектора та центральної послідовності, що утворюються після обробки вектора рестриктазами були не комплементарними. З генетичної карти вектора видно, що після двох послідовних обробок рестриктазами EcoRI та BamHI „ липкі” кінці плеч вектора утворені рестриктазою BamHI, а липкі кінці центральної частини рестриктазою EcoRI. „Липкі” кінці фрагменту ДНК мають бути комплементарними кінцям «плеч» вектора.
3. Гормон інсулін складається з двох поліпептидних ланцюгів, перший – з 21 амінокислотного залишку (А-ланцюг), другий – з 30 (В-ланцюг), з’єднаних дисульфідними зв’язками. Хромосомний ген кодує проінсулін, в якому ці два ланцюги зв’язує між собою петлеподібний пептид, який видаляється під дією ферментів під час визрівання в везикулах підшлункової залози. До того ж ген містить інтрон. Яким чином можна одержати інсулін в клітинах E.coli?
Розв’язання.
Необхідно використати вектор, який може реплікуватись в клітинах E.coli. Оскільки довжина фрагменту, що клонується, невелика, можна обрати як вектор плазміду. В складі такої плазміди обов’язково повинні бути: ділянка ori та сильний регульований промотор (наприклад, β-галактозидази). Оскільки, ген проінсуліну містить інтрон, необхідно з допомогою зворотної транскрипції за матрицею м-РНК створити кодуючи ДНК. Кодуючи ДНК необхідно розмістити за промотором β-галактозидази та ініціюючим кодоном ATG, що знаходиться на певній відстані за промотором та ділянкою Шайна-Дальгарно. Це можна зробити без збереження рамки зчитування, тоді утвориться химерна ДНК та химерний білок. Можна розмістити ген проінсуліну після гену β-галактозидази через метіоніновий кодон із збереженням рамки зчитування. Тоді утвориться гібридна ДНК та гібридний білок. Обов’язковою умовою є наявність сигналів термінації одразу після структурної ділянки гену. Вищевказані операції проводяться методами рекомбінантних ДНК (дія рестриктаз, приєднання до кодуючої ДНК „липких” кінців, зшивання з допомогою лігази, скринінг клонотеки генів та інше). Від химерного або гібридного білку проінсулін можна відділити хімічними методами, а зробити з проінсуліну інсулін можна з допомогою дії ферментів.
Примітка. В промисловості ДНК, що кодує інсулін, часто синтезується хімічно, а в якості промотору використовується промотор гену β-лактамази (гену стійкості до ампіциліну) з утворенням гібридного білку, що забезпечує секрецію проінсуліну з бактеріальних клітин завдяки присутності сигнального пептиду на N-кінці β-лактамази.
4. Фермент SOD людини використовується для лікування запальних процесів та зменшення побічної дії від введення інших ферментів. Попередні досліди з клонування к-ДНК проведені в E.coli, але якість одержаного білку була недостатньою у зв’язку з відсутністю сигналів посттрансляційної модифікації. Тому для одержання якісного ферменту к-ДНК була введена в склад експресуючого бінарного вектору для S. cerevisiae. Зобразіть генетичну карту такого вектору, покажіть необхідні ділянки.
Р озв’язання.
озв’язання.
Дріжджовий бінарний експресуючий вектор обов’язково містить такі ділянки: точку ori та селективний маркер для E.coli (наприклад, ген Аmp), точку ori та послідовності, що забезпечують стабільність позахромосомного існування для S.cerevisiae (S1, S2), дріжджовий селективний маркер (наприклад, ген лейцину Leu), промотор дріжджового гену (G, гліцеральдегідрофосфатдегідрогінази, цей промотор нерегульований, тому синтез ферменту продовжується постійно). К-ДНК розміщена між промотором G та сигналами термінації та поліаденілірування Gp. Всі попередні процедури проводять в E.coli, після чого цим штамом трансформують штам дріжджів, не здатний до синтезу лейцину.
5. Вам необхідно синтезувати поверхневий антиген вірусного гепатиту В в дріжджах Pichia pastoris, використовуючи інтегруючий експресуючий вектор, якщо всі попередні маніпуляції проводяться в E.coli.
Розв’язання.
І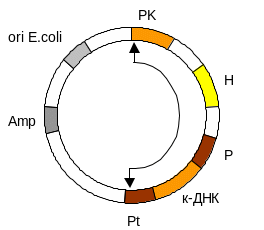 нтегруючий
вектор для одержання експресії в клітинах
P.pastoris
обов’язково містить: сайт ініціації
реплікації та селективний маркер
(наприклад, Amp)
для E.coli;
послідовності, комплементарні певним
хромосомним послідовностям P.pastoris
(PK),
між якими розміщується чужорідний ген,
експресія якого очікується . Чужорідний
ген знаходиться за промотором
дріжджового
гену (Р)
(наприклад, алкогольоксидази) безпосередньо
перед сигнальними послідовностями
термінації–поліаденілірування цього
гену (Pt).
Інтегрована
ділянка містить селективний дріжджовий
маркер (Н), який робить можливим ріст
клітин на середовищі без гістаміну.
Операції клонування проводяться в
E.coli.
нтегруючий
вектор для одержання експресії в клітинах
P.pastoris
обов’язково містить: сайт ініціації
реплікації та селективний маркер
(наприклад, Amp)
для E.coli;
послідовності, комплементарні певним
хромосомним послідовностям P.pastoris
(PK),
між якими розміщується чужорідний ген,
експресія якого очікується . Чужорідний
ген знаходиться за промотором
дріжджового
гену (Р)
(наприклад, алкогольоксидази) безпосередньо
перед сигнальними послідовностями
термінації–поліаденілірування цього
гену (Pt).
Інтегрована
ділянка містить селективний дріжджовий
маркер (Н), який робить можливим ріст
клітин на середовищі без гістаміну.
Операції клонування проводяться в
E.coli.
Примітка. Вектор , що використовується у виробництві містить також ділянку ori для забезпечення реплікації в P.pastoris .
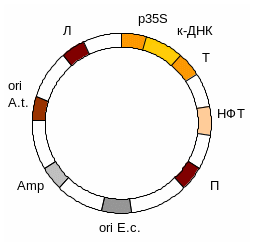
6. Вам необхідно створити трансгенну рослину тютюну, стійку до дії певного гербіциду, застосовуючи коінтегративний вектор на основі Ті-плазміди. Відомо, що стійкість рослини до цього гербіциду забезпечується трансформацією рослини тютюну бактеріальним геном нітрилази, яка нейтралізує дію гербіциду.
Розв’язання.
Коінтегративний вектор містить сайт ініціації реплікації та селективний маркер (наприклад, Amp) для E.coli, праву фланкуючу послідовність Т-ДНК Ті-плазміди, рослинний селективний маркер (наприклад, ген стійкості до канаміцину - Kan) , ген нітрилази під контролем сильного рослинного промотору (наприклад вірусу кольорової мозаїки), після якого обов’язково знаходяться сигнали термінації та поліаденілірування . Цей вектор також обов’язково містить послідовність. гомологічну ділянці допоміжної неонкогенної Ті-плазміди для забезпечення їх інтеграції. Допоміжна плазміда містить сайт ініціації реплікації в A.tumefaciens та гени vir а також ліву фланкуючу послідовність Т-ДНК. Коінтегративна плазміда трансформує клітини E.coli, після чого переноситься шляхом кон’югації в реціпієнтний штам A.tumefaciens. Після рекомбінації ген токсину входить до складу хромосоми тютюну.
В
 ам
необхідно створити трансгенну рослину
томату, стійку до шкідливих комах,
використовуючи бінарний вектор на
основі Ті-плазміди. Відомо, що з одного
з видів бацил виділили ген природного
токсину. Зобразіть генетичну карту
цього вектору.
ам
необхідно створити трансгенну рослину
томату, стійку до шкідливих комах,
використовуючи бінарний вектор на
основі Ті-плазміди. Відомо, що з одного
з видів бацил виділили ген природного
токсину. Зобразіть генетичну карту
цього вектору.
Розв’язання.
Бінарний вектор на основі Ті-плазміди містить сайт ініціації реплікації та селективний маркер (наприклад, Amp) для E.coli, між лівою та правою фланкуючими послідовностями знаходяться : рослинний селективний маркер (наприклад, неоміцинфосфотрансфераза) та ген природного токсину під контролем сильного рослинного промотору (наприклад вірусу кольорової мозаїки), після якого обов’язково знаходяться рослинні сигнальні послідовності термінації та поліаденілірування. Цей вектор вводять в штам A.tumefaciens, яким заражують рослини томату.
8. Пізню ділянку вірусу SV40 замінили на ген α-глобіну миші і одержали змішану популяцію вірусів, в якій були присутні рекомбінантні молекули ДНК вірусу SV40. Які ще вірусні частинки присутні в цій популяції?
Розв’язання.
Вектор із заміщенням пізньої ділянки геному використовують разом з допоміжним вірусом з дефектною ділянкою ранніх білків. Допоміжний вірус продукує білки капсиду . В результаті утворюється змішана популяція, що складається з вірусних частинок SV40 з дефектною ранньою ділянкою та частинок SV40, що містять глобіновий ген.
9. Ранню ділянку вірусу SV40 замінили на ген гематглютину вірусу грипу та одержали рекомбінантні молекули, що спакувались у віріони. Яку клітинну культуру використовували в цьому випадку для трансфекції?
Розв’язання.
Вектор із заміщенням ранньої ділянки геному трансфекує спеціально підготовлені клітини COS, одержані в результаті зараження клітин зеленої мавпи вірусом SV40 з нормальними послідовностями , що кодують ранні та пізні білки, але дефектною ділянкою ori . В результаті вірусна ДНК інтегрує з хромосомами клітин, де продовжує продукувати Т-антиген, гену якого не вистачає в рекомбінантній вірусній молекулі.
10. Ген тимідинкінази вірусу герпесу клонували в векторній молекулі, що містить сайт ініціації реплікації SV40 та трансфекували клітини миші. Зливання цих клітин з клітинами COS ініціює утворення (в частині випадків) кільцевих молекул. Які ділянки повинні бути присутні в вихідній векторній молекулі, щоб одержані кільцеві молекули можна було розмножити та ідентифікувати в E.coli?
Розв’язання.
Очевидно, одержана кільцева молекула ампліфікується в E.coli у вигляді плазміди. Для цього їй необхідний сайт ініціації реплікації ori E.coli, таким чином вектор має бути бінарним. Для ідентифікації кільцевої молекули в E.coli в ній має бути відповідний селективний маркер (наприклад, Amp).
11. Для одержання експресії генів субодиніць α та β димерного білку в клітині ссавця було використано двоцисторний експресуючий вектор. Покажіть на генетичній карті вектора положення генів α та β, зазначте, які ще послідовності необхідні в складі такого вектору.
Розв’язання.
Двоцисторний вектор обвязково містить сайт ініціації реплікації та селективний маркер для E.coli, сайт ініціації реплікації та селективний маркер для клітини ссавця ,гени субодиниць білків α та β розміщені під контролем еукаріотичного промотора та сигналу поліаденілірування, між ними із збереженням рамки зчитування та сигналів ініціації розташована зв’язуюча послідовність ДНК.
12. Онкогенний вірус використали як вектор для доставки терапевтичного гену С в клітини потрібної тканини так, як показано на генетичній карті :
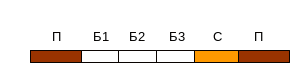
В якому випадку необхідно остерігатись злоякісної трансформації цих клітин?
Розв’язання.
Як ми бачимо, в самому векторі онкогену нема, відповідно цей вірус не трансформує клітини, але може активувати клітинний онкоген внаслідок інтеграції в близько розміщену ділянку ДНК.
13. Екзогенна ДНК вбудувана в склад ретровірусного вектору так, як це показано на генетичній карті, інтегрувала в профаг. Трансляція яких білків відбудеться з цієї ділянки?
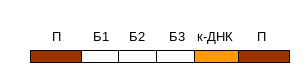
Розв’язання.
При транскрипції ДНК інтегрованого провірусу утворюється первинний транскрипт. Відбувається його диференційований сплайсинг. Можливі альтернативні шляхи сплайсінгу цього транскрипту з утворенням м-РНК для різних білків:
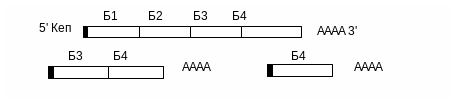
Таким чином синтезуються наступні білки: Б1, Б2, Б3 , в тому числі і білок, синтез якого ми очікували.