

3.25.3. Организация манипуляторных движений
Манипуляторные движения — это наиболее яркий пример произвольных движений, возникновение которых обусловливают внутренние мотивационные влияния. Эти движения являются локальными, что ставит перед двигательной системой следующие задачи: 1) выбор ведущего мышечного звена; 2) компенсацию внешней нагрузки; 3) настройку позы; 4) соотнесение координат цели и положения собственного тела.
Отличительной чертой манипуляторных движений является их зависимость от центральной программы и телецептивных сенсорных входов. Поэтому ведущая роль отводится следующим мозговым структурам: лобной коре, базальным ядрам и мозжечку. Активность других супраспинальных цепей двигательного контроля в большей степени зависит от периферической информации, поступающей вслед за реализацией команд.
Ведущая роль в программировании быстрых движений принадлежит мозжечковой системе, а в программировании медленных — базальным ядрам. Выходы этих структур на спинальный уровень опосредованы в основном через сенсомоторную кору, которая дополнительно к этому обладает своими собственными функциями.
3.25.4. Корковая сенсомоторная интеграция
Структурной единицей неокортекса является модуль — микросистема тесно контактирующих между собой пирамидных нейронов для их синхронного вовлечения в формирование эфферентного импульсного залпа (рис. 3.68). Такое объединение рассматривают в качестве жесткого структурного элемента организации коры, формирующегося в пренатальном онтогенезе. Почти всем модулям свойственна широкая конвергенция гетеросенсорных влияний. В моторную кору конвергируют те компоненты сенсорной информации, которые являются жизненно важными для организма. Это информация о биологической значимости того или иного сигнала либо его значимости, приобретенной в порядке выработки условного рефлекса.
Топографическое распределение соматических проекций участков тела на кору больших полушарий только частично определяется анатомическими закономерностями. Не менее важную роль играют функциональные механизмы, определяющим среди которых является внутрикорковое торможение.
Структурный корковый модуль выступает в качестве элементарной сенсо—моторной единицы. Различия между отдельными модулями определяются характером соматической сенсорной проекции и адресатом эфферентного выхода из того же модуля. Существует распределенная система корковых модулей, выходы которых конвергируют на разных уровнях ствола и спинного мозга, обеспечивая управление деятельностью определенной мышечной группы. Распределенная система является частью более обширных общемозговых систем, сформированных в раннем онтогенезе. Такие функциональные структуры управляют более обширной частью скелетно—мышечного аппарата.
|
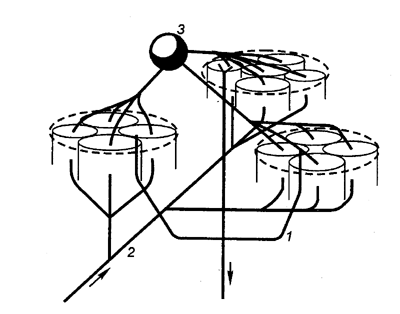
Рис. 3.68 Функциональные объединения нейронных модулей Одиночный структурный модуль имеет вид цилиндра; 4—6 модулей могут сгруппироваться в функциональные концентрические объединения, между которыми . возможны как облегчающие, так и взаимотормозные отношения. Обширные функциональные объединения из нескольких концентрических модулей строятся на основе ветвлений аксонной коллатерали пирамидных нейронов (1), ветвлений таламического афферента (2) и ветвлений аксона корзинчатой клетки (3); стрелками показано направление восходящей и нисходящей импульсации. |
Вовлечение корковых систем в целостную деятельность мозга определяется текущей мотивацией и конкретной двигательной программой, направленной на удовлетворение этой мотивации. Двигательная программа, чтобы носить приспособительный характер, должна учитывать все сигнально значимые компоненты внешней среды, относительно которых совершается целенаправленное движение, т. е. строиться на принципе мультисенсорной конвергенции. Телецептивные входы на модулях сенсомоторной коры носят сравнительно диффузный характер и адресуются ко многим пространственно разнесенным одноименным модулям. Констелляции модулей основаны на синергичных и антагонистичных межмодульных соотношениях, что обеспечивает динамичность их участия в различных актах сенсомоторной интеграции.