

Морфо-функциональная характеристика иммунной системы
Общая морфо-функциональная характеристика иммунной системы
Иммунная система осуществляет защиту организма от генетически чужеродных тел и веществ, называемых антигенами. Такая функция иммунной системы обеспечивается ее способностью отличать "свое" от "чужого" и обеспечивать ингибирование и элиминацию генетически чужеродных структур. Причем под генетически чужеродными структурами понимают не только посторонние организмы (вирусы, бактерии) и синтезированные ими биополимеры, но и собственные генетически измененные в результате мутаций при воспроизведении клетки организма. Следовательно, основной физиологической функцией иммунной системы является специфическая иммунологическая реактивность, реализующаяся благодаря тому, что эта система
концентрирует антиген
обеспечивает контакт с антигеном различных видов клеток
транспортирует гуморальные вещества и клеточные структуры лимфоидной ткани в необходимые участки организма
в конечном итоге элиминирует чужеродный антиген.
Иммунологическая реактивность может проявляться в виде двух типов процессов:
выработке антител, которые либо непосредственно уничтожают антиген, либо облегчают его уничтожение макрофагами, нейтрофилами или системой комплимента (гуморальные иммунные реакции)
в непосредственном повреждающем действии лимфоцитов по отношению к антигенам (клеточные иммунные реакции).
Для иммунной системы характерны следующие морфо-функциональные особенности:
она генерализована по всему телу
ее клетки постоянно рециркулируют по всему организму через лимфо- и кровоток
она обладает уникальной способностью вырабатывать сугубо специфические молекулы антител, различные по своей специфике в отношении каждого антигена.
В организме человека содержится примерно 1011 делящихся клеток. Если допустить, что мутации возникают в 10-5 – 10-7 случаев делений (подобно бактериям), то в организме человека при однократном воспроизведении пула пролиферирующих клеток должно образовываться примерно 105-106 мутантных клеток. В норме такие клетки или возникшие из них конгломераты ингибируются иммунной системой и элиминируются. Таким образом, иммунная система обеспечивает поддержание генетического гомеостаза в организме, а, следовательно, гомеостаза физико-химической структуры макромолекул и клеток. Дефекты специфической иммунологической реактивности приводят либо к потере способности различать "чужое" (вследствие чего может возникнуть рак и длительные инфекционные заболевания), либо утрате способности различать "свое" (что служит причиной возникновения аутоиммунных заболеваний вследствие реакции иммунной системы против собственных тканей).
Наряду со специфической иммунологической реактивностью иммунной системы, проявляющейся в ее способности распознавать "чужое" и обеспечивать нетерпимость к нему, она обладает не менее важным свойством иммунологической толерантности, заключающейся в способности обеспечивать терпимость к "своему".
Органы иммунной системы расположены в различных частях организма и анатомически обособлены друг от друга; их основа образована лимфоидной или миелоидной тканью. Миелоидная ткань (от греч. миелос – костный мозг) располагается в мозгу, находящемся в полостях костей (костном мозгу), и является местом образования эритроцитов, кровяных пластинок, гранулоцитов, моноцитов и В-лимфоцитов (т.е. почти всех форменных элементов крови за исключением Т-лимфоцитов). Лимфоидная ткань получила свое название в связи с очень большим изобилием лимфоцитов, в ней находящихся; она образует тимус, селезенку, лимфатические узлы и неинкапсулированные лимфоидные скопления в рыхлой волокнистой соединительной ткани слизистых оболочек и кожи.
В кроветворных органах (красном костном мозге, образованном миелоидной тканью, и тимусе, образованном лимфоидной тканью) в ячеях ретикулярной ткани расположены созревающие клетки крови и их предшественники.
В основе лимфоидных органов лежит ретикулярная паренхима, образованная в большинстве случаев ретикулярной тканью (а в тимусе – ретикулярной и эпителиальной тканью) и имеющая вид сети, в ячеях которой находятся лимфоциты, являющиеся главными функциональными элементами лимфоидной ткани. Лимфоидные органы составляют 5% от массы тела человека (общая их масса составляет в среднем 1,5-2 кг), а общее число лимфоидных клеток – 1012. Лимфоциты (Т- и В-лимфоциты) представляют собой иммунологически компетентные клетки, служащие ключевым звеном:
в реакциях гиперчувствительности замедленного типа и трансплантационного иммунитета (формы клеточного иммунитета),
предшественниками антителообразующих клеток (плазматических клеток), обеспечивающих осуществление гуморальных иммунных реакций
носителями иммунологической памяти
кроме того, лимфоциты выделяют ряд гуморальных факторов, вовлекающих в иммунный ответ иммунокомпетентные и вспомогательные клетки, вступают в кооперативное взаимодействие с ними.
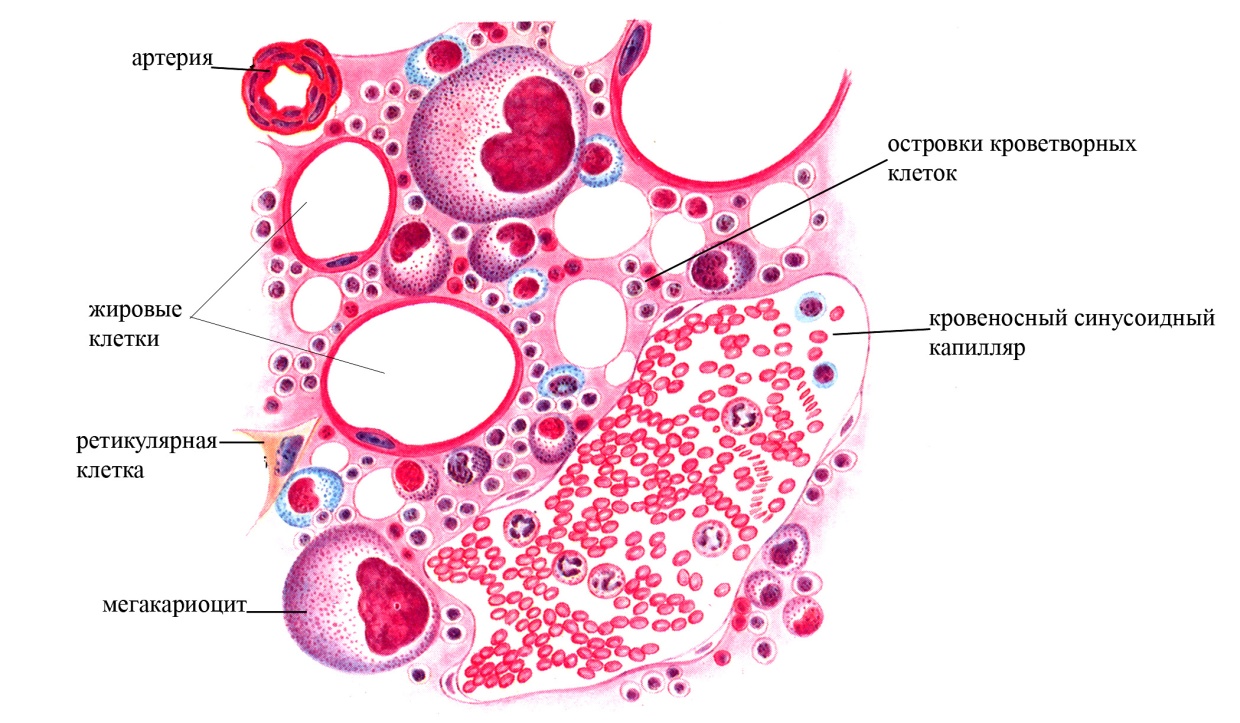
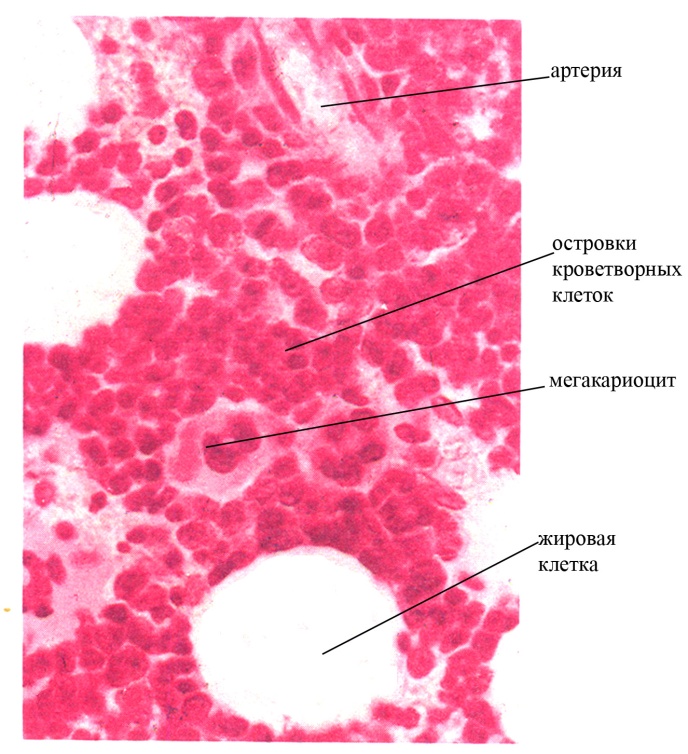 А
А
Б
Рис. 1 Красный костный мозг. А – схема строения. Б – микрофотография красного костного мозга (окраска гематоксилин-эозином)
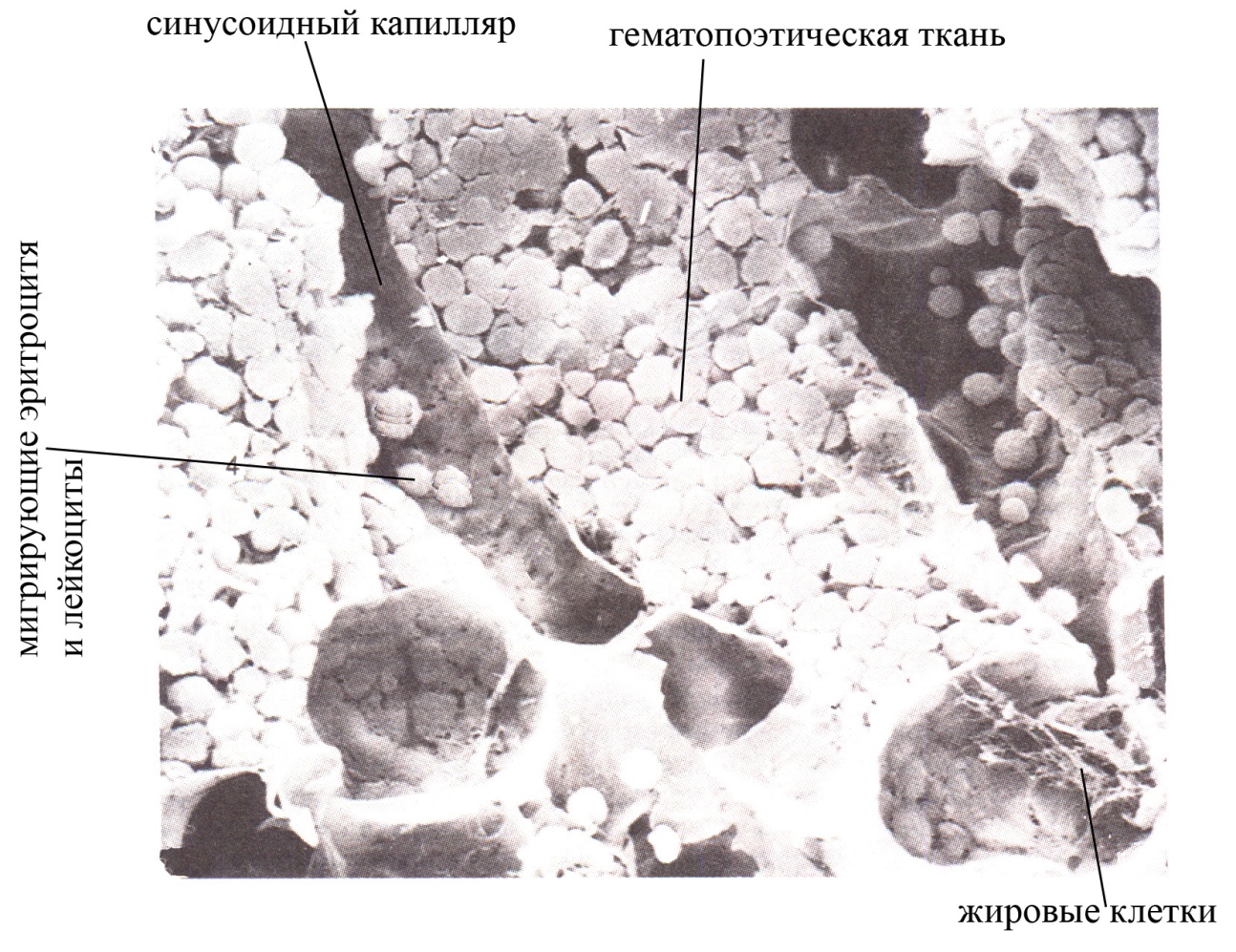
Рис. 2. Синусоидный капилляр красного костного мозга (вид при сканирующей электронной микроскопии)
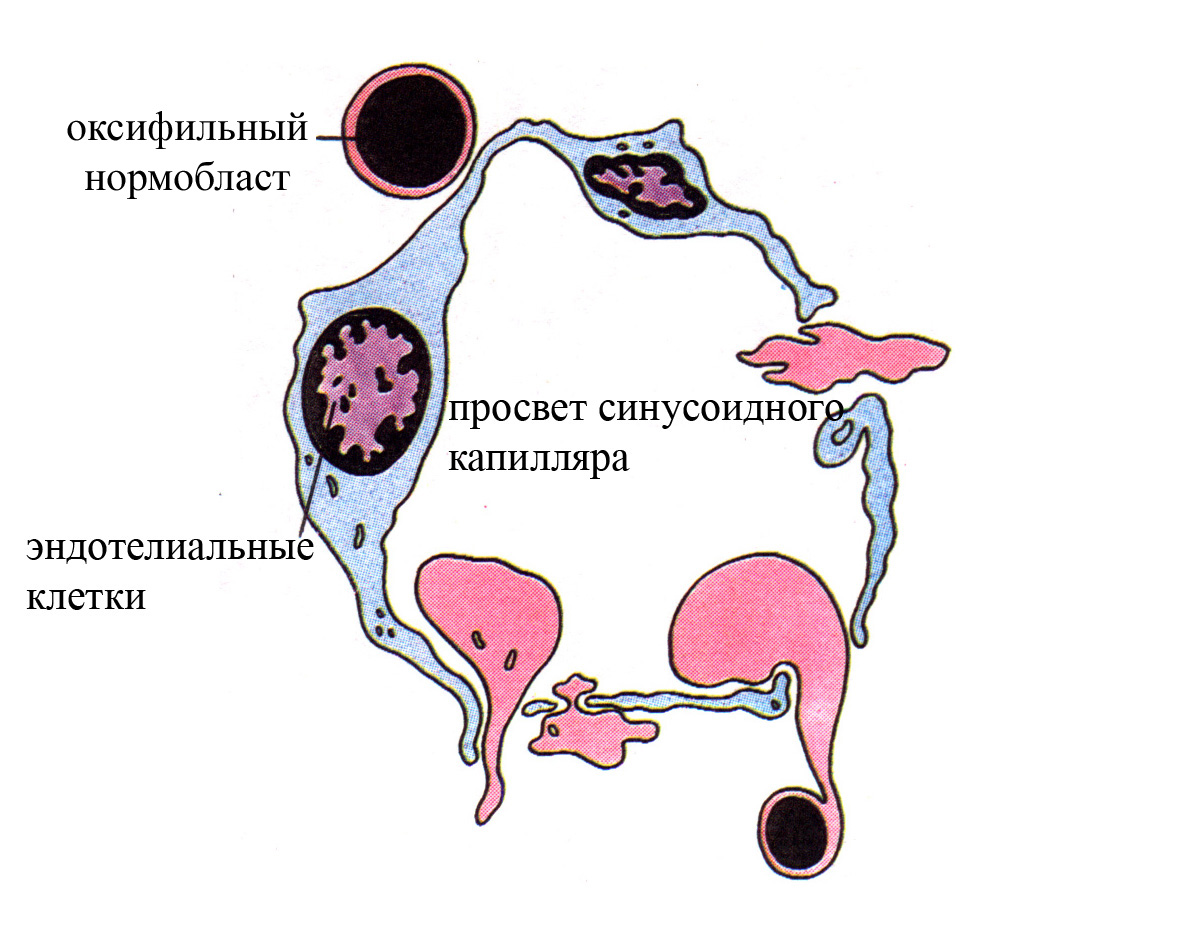
Рис. 3. Схема, демонстрирующая выход эритроцитов через стенку синусоидного капилляра красного костного мозга в кровоток
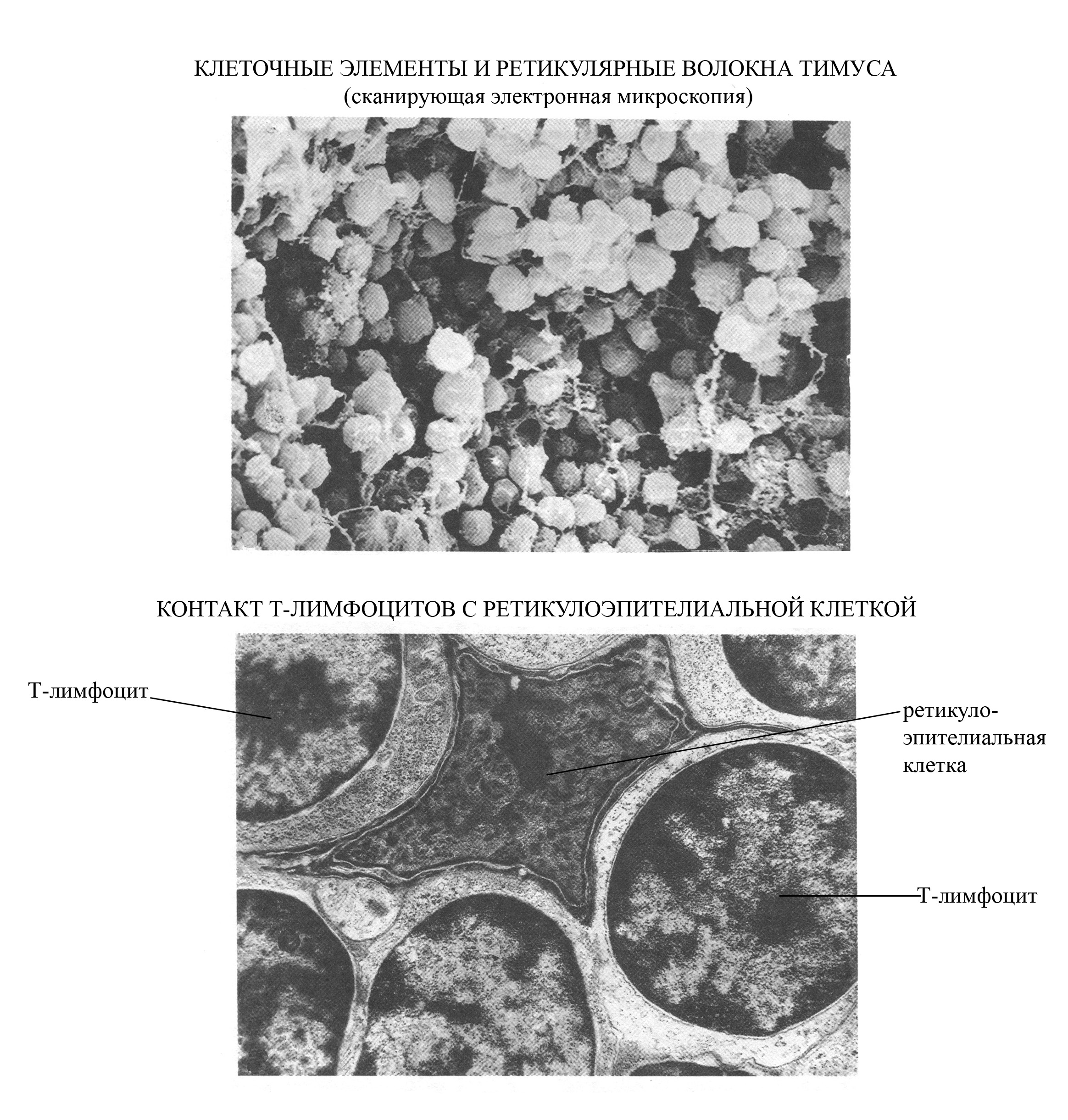
Рис. 4. Эпителиальная ткань тимуса (вид при сканирующей электронной микроскопии)
Органы иммунной системы в зависимости от их функционального назначения классифицируют на:
центральные органы иммунной системы – органы, в которых осуществляется лимфоцитопоэз, служат своеобразной "фабрикой" по производству лимфоцитов. Следовательно, центральные органы иммунной системы, являясь местом лимфоцитопоэза, сообщают иммунологическую компетентность клеткам-предшественникам и регулируют иммунологическую реактивность всей иммунной системы посредством вырабатываемых ими гуморальных факторов. Причем сами эти органы не осуществляют активного иммунного ответа, они заселяются стволовыми клетками и являются местом усиленного лимфопоэза даже в эмбриональном периоде и у безмикробных животных, когда периферические лимфоидные органы остаются неактивными. Удаление центральных органов иммунной системы приводит к нарушению функций иммунитета, резкому снижению числа лимфоцитов в периферической крови и периферических лимфоидных органах. Эти органы представлены:
красным костным мозгом (образован миелоидной тканью). У взрослого человека располагается в полостях губчатого костного вещества костей свода черепа, ребер и грудины, телах позвонков, губчатых частях некоторых коротких костей и в эпифизах длинных трубчатых костей. Наряду с эритроцитами, кровяными пластинками, гранулоцитами и моноцитами, служит местом образования В-лимфоцитов. Причем на долю лимфоцитов в красном костном мозге приходится 10% от всех ядерных его клеток. Кроветворение именно в красном костном мозге обусловлено тем, что он заложен в полостях костной ткани, которые не могут бесконечно расширяться (в отличие, например, от волокнистых соединительных тканей), и это заставляет образующиеся зрелые форменные элементы крови переходить в капилляры. Причем для костного мозга характерны расширенные капилляры с прерывистым эндотелием (синусоидные капилляры), что облегчает переход в них эритроцитов и других форменных элементов. Сами же синусоидные капилляры костного мозга не могут спадаться под действием окружающих их форменных элементов благодаря высокому гидростатическому давлению в них, обусловленному тем, что приносящие артериолы, ответвляющиеся от костных артерий, шире венул. Такая особенность микроциркуляторного русла, обеспечивающая повышенное гидростатическое давление в капиллярах, в других местах организма приводила бы к отеку, который не возникает в костной ткани благодаря большой плотности кости.
тимусом (вилочковая железа), лимфоидный орган, является местом образования Т-лимфоцитов. Причем все форменные элементы крови происходят от общего родоначального предшественника – стволовой клетки (называемой также колониеобразующей единицей – КОЕ). Эта стволовая клетка или, возможно, ее потомки с током крови из красного костного мозга приносятся в тимус, где дают начало Т-лимфоцитам.
периферические органы иммунной системы – органы, в которых в отличие от центральных органов иммунной системы не происходит первичного лимфоцитопоэза (т.е. антигеннезависимого лимфоцитопоэза из кроветворных стволовых клеток). Периферические органы иммунной системы заселяются лимфоцитами и служат местом контакта лимфоцитов с антигенами, т.е. представляют собой плацдарм для осуществления иммунных реакций (преимущественно гуморального типа), в результате которых происходит элиминация антигенов. К периферическим органам иммунной системы относят:
лимфатические узлы, где осуществляется фильтрация лимфы, оттекающей из тканей и ее очищение от различных антигенов (бактерий, вирусов, их белковых макромолекул). Лимфа образуется в тканях путем фильтрации тканевой жидкости в слепо начинающиеся в тканях лимфатические капилляры. Причем особенностью лимфатических капилляров, в отличие от кровеносных, является отсутствие базальной мембраны, что облегчает прохождение в них крупномолекулярных белков, воспалительных экссудатов и даже отдельных клеток из тканей. Лимфатические капилляры сливаются друг с другом, переходя в более крупные сосуды, выносящие лимфу из органов. Лимфатические сосуды, прежде чем доставить лимфу в кровоток, обязательно проходят через лимфатические узлы. Причем они входят в лимфатические узлы со стороны выпуклой их поверхности, а выходят – со стороны вогнутой. Лимфа, выходящая из лимфатического узла, обогащена лимфоцитами (в основном Т-лимфоцитами) и частично очищена от антигенов. Лимфатические сосуды, прошедшие через лимфатические узлы, по ходу сливаются друг с другом, образуя все более крупные сосуды. Наконец, лимфа через грудной и правый лимфатические протоки впадает в верхнюю полую вену (возвращается в кровь). Таким образом, система лимфообращения непрерывно выносит лимфу в венозную часть системы кровообращения, так что вся избыточная жидкость, формирующаяся в организме, возвращается в кровяное русло, проходя на своем пути лимфатические узлы.
селезенка, выступает в качестве депо крови, принимая участие в очистке ее от различных антигенов, обеспечивает элиминацию старых эритроцитов
неинкапсулированная лимфоидная ткань, заложенная в рыхлой соединительной ткани слизистых оболочек и кожи, обеспечивает очистку от антигенов преимущественно тканевой жидкости.
Р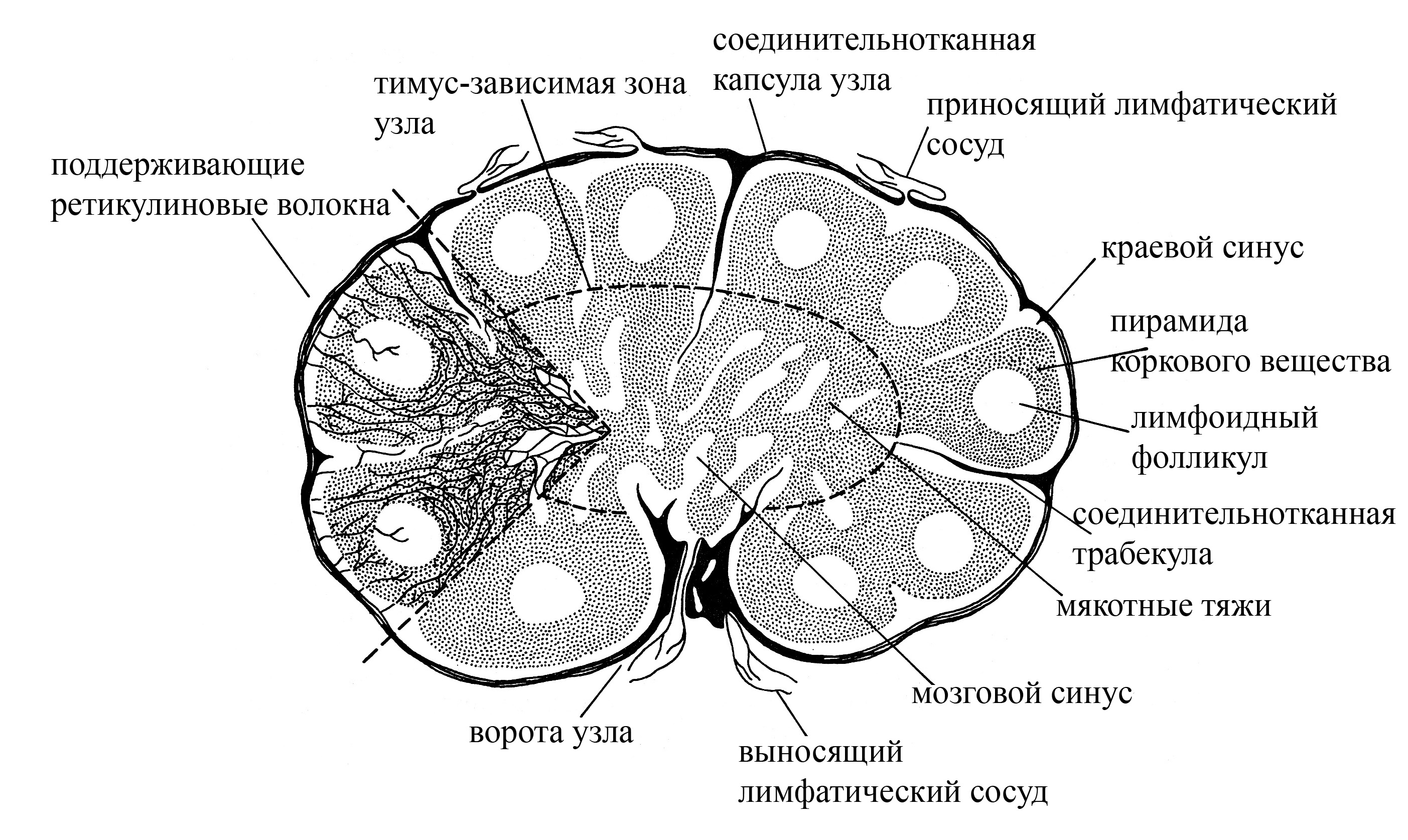 ис.
5. Лимфатические узлы и сосуды головы и
шеи человека
ис.
5. Лимфатические узлы и сосуды головы и
шеи человека
Рис. 6. Схема строения лимфатического узла. Обратите внимание на то, что приносящие лимфатические сосуды входят в лимфатический узел со стороны наружной (выпуклой) его поверхности. После чего лимфа изливается в краевой лимфатический синус, расположенный под соединительнотканной капсулой узла. Из краевого лимфатического синуса лимфа поступает в промежуточные синусы коркового вещества, откуда переходит в мозговые синусы, открывающиеся в выносящие лимфатические сосуды, которые выходят из узла в области ворот. Таким образом, лимфа фильтруется через лимфатический узел и, проходя по его синусам, постоянно контактирует с лимфоцитами и макрофагами, находящимися в лимфатических синусах, что обеспечивает очистку лимфы от различных антигенов.
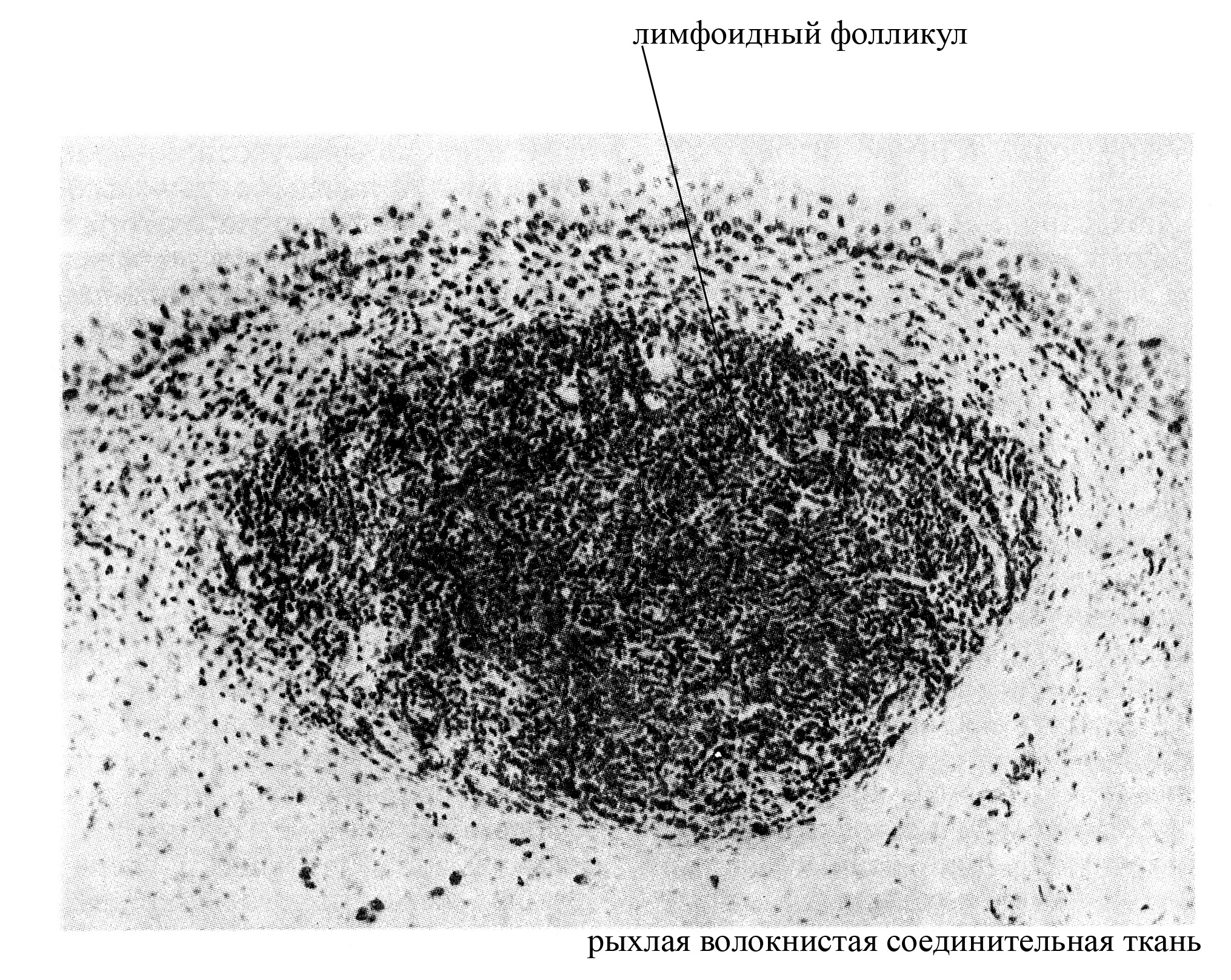
Рис. 7. Неинкапсулированный лимфоидный фолликул рыхлой волокнистой соединительной ткани
Лимфатические узлы и неинкапсулированные лимфоидные скопления расположены в большинстве своем на путях проникновения инфекционных агентов в организм (в области кожи, слизистых оболочек), а, следовательно, выполняют защитно-барьерную функцию. Причем между периферическими органами иммунной системы существует "своеобразное разделение труда": они специализируется на контакте с антигенами определенных компонентов внутренней среды организма. Так, неинкапсулированные лимфоидные фолликулы контактируют преимущественно с антигенами, которые оказываются в тканевой жидкости, лимфатические узлы эффективно взаимодействуют с лимфой, протекающей через них. Наконец, в селезенке осуществляется активный и весьма длительный контакт иммуннокомпетентных клеток с антигенами, находящимися в крови, протекающей через селезенку.
Особенностью иммунной системы является ее адаптивная направленность, позволяющая обеспечить иммунный ответ на любые теоретически возможные чужеродные макромолекулы и клетки, которые могли бы проникнуть во внутреннюю среду организма. Данное свойство иммунной системы обусловлено тем, что значительное количество клеток, отвечающих специфически на данный антиген, образуется только после специфического антигенного стимула (т.е. после контакта с каким-то антигеном) и, будучи индуцированы, эти клетки выполняют свои специфические функции автономно. Следовательно, приобретение лимфоцитами иммунокомпетентности происходит не в центральных иммунных органах, где осуществляется лимфоцитопоэз, а в периферических иммунных органах после контакта с теми или иными антигенами.
Иммунная система по организации и механизмам функционирования сходна с нервной системой. Сходство заключается в том, что функции обеих этих систем генетически детерминированы, обе они представлены центральными и периферическими органами, способны реагировать на различные сигналы, имеют огромное количество рецепторных структур, специфическую память. Основным же функциональным отличием иммунной системы от нервной является то, что иммунная система способна к передаче и генерализации специфических сигналов мигрирующим клеткам (лимфоцитам), тогда как нервная – неподвижным клеткам.
Морфо-функциональная характеристика тимуса
Тимус (вилочковая железа) представляет собой лимфоидный орган, расположенный большей частью в грудной клетке, непосредственно позади рукоятки грудины, в межплевральной области, спереди перикарда. Тимус состоит из двух долей, соединенных между собой и образующих единое целое, имеет удлиненную уплощенную треугольную форму, розовато-серый цвет, вершина его направлена в область шеи.
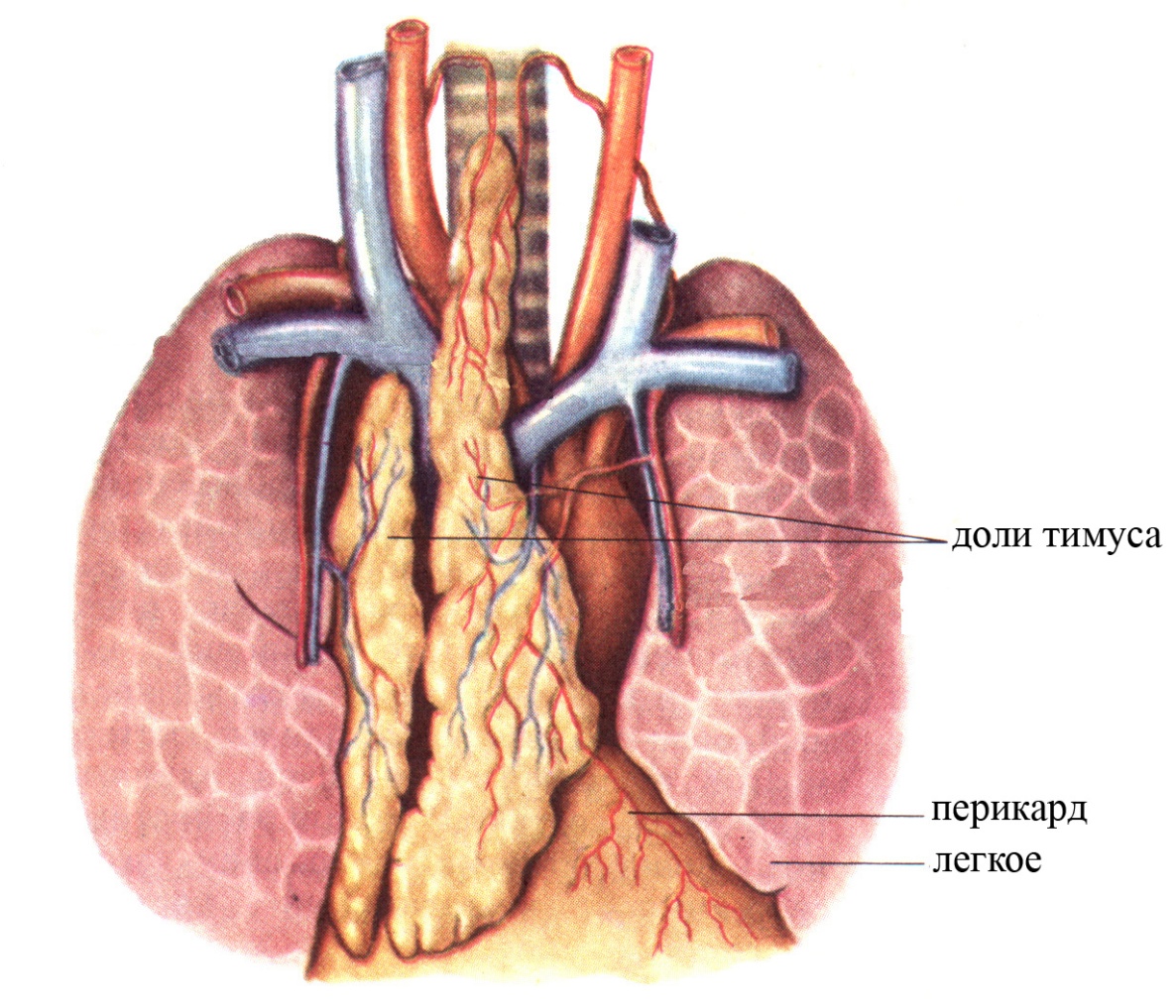
Рис. 8. Тимус (общий вид).
Вилочковая железа энтодермального происхождения. Ее эпителиальный компонент начинает закладываться в конце 1-го - начале 2-го месяца внутриутробной жизни из 3-го глоточного кармана в виде парных тяжей, которые в дальнейшем утрачивают связь с глоткой, опускаются в грудную полость и срастаются между собой. Мезенхима, окружающая эпителиальные зачатки тимуса, дает начало его соединительнотканной капсуле и многочисленным ее производным – септам, формирующим в совокупности соединительнотканную строму органа. Причем соединительнотканные септы тимуса, отходящие от его капсулы, не обеспечивают полного разделения паренхимы тимуса на дольки (орган имеет псевдодольчатое строение). Вскоре между эпителиальными клетками тимуса начинают появляться лимфоциты, которые дифференцируются либо из самой кроветворной стволовой клетки либо из ее потомков – предшественников лимфоцитов, мигрирующих из красного костного мозга. Лимфоциты становятся главными клетками тимуса. Таким образом, в тимусе можно выделить три гистологических компонента:
эпителиальный, являющийся производным энтодермы глоточного кармана,
соединительнотканный, имеющий мезенхимное происхождение и формирующий соединительнотканную капсулу и строму органа,
лимфоидный, представленный лимфоидными клетками на разных стадиях дифференцировки, которые являются производными стволовой кроветворной клетки, мигрирующей из красного костного мозга.
Тимус формируется раньше других органов иммунной системы и к моменту рождения имеет значительную массу (10-15 г), в течение первых 3-ех лет постнатальной жизни тимус растет наиболее интенсивно, а затем его масса и размеры относительно стабилизируются (достигая 30-40 г) и остаются таковыми до окончания периода пубертации (в среднем до 20 лет), после чего начинается возрастная инволюция тимуса (в пожилом возрасте масса органа уменьшается до 13-15 г). Причем возрастная инволюция тимуса сопровождается не просто уменьшением его массы, но и определенными морфологическими изменениями. В частности, в органе увеличивается доля соединительной и жировой ткани (у людей старше 50 лет она составляет 90% от массы органа), значительно сокращается объем эпителиальной паренхимы и лимфоидного компонента.
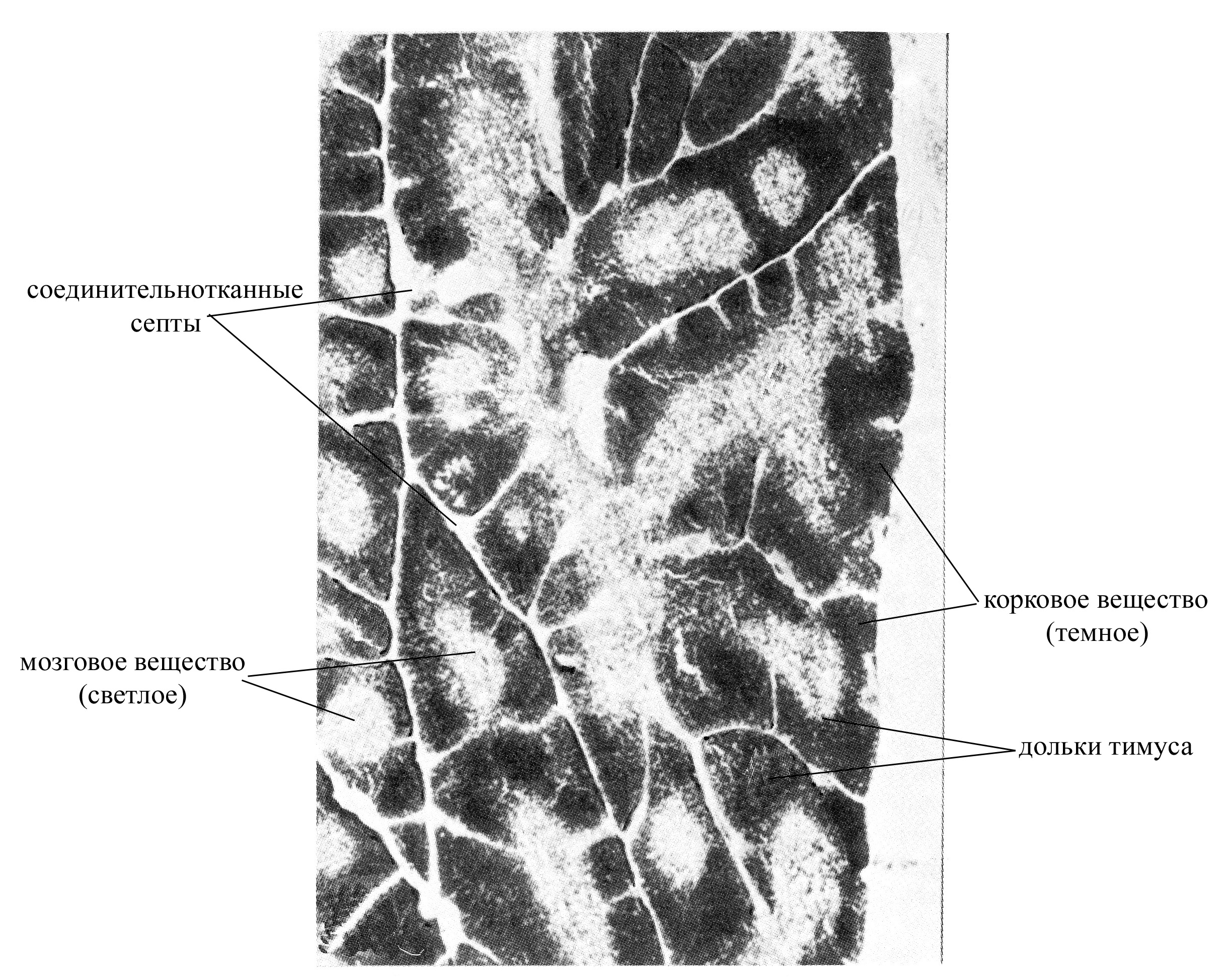
Рис. 9. Микрофотография тимуса при малом увеличении
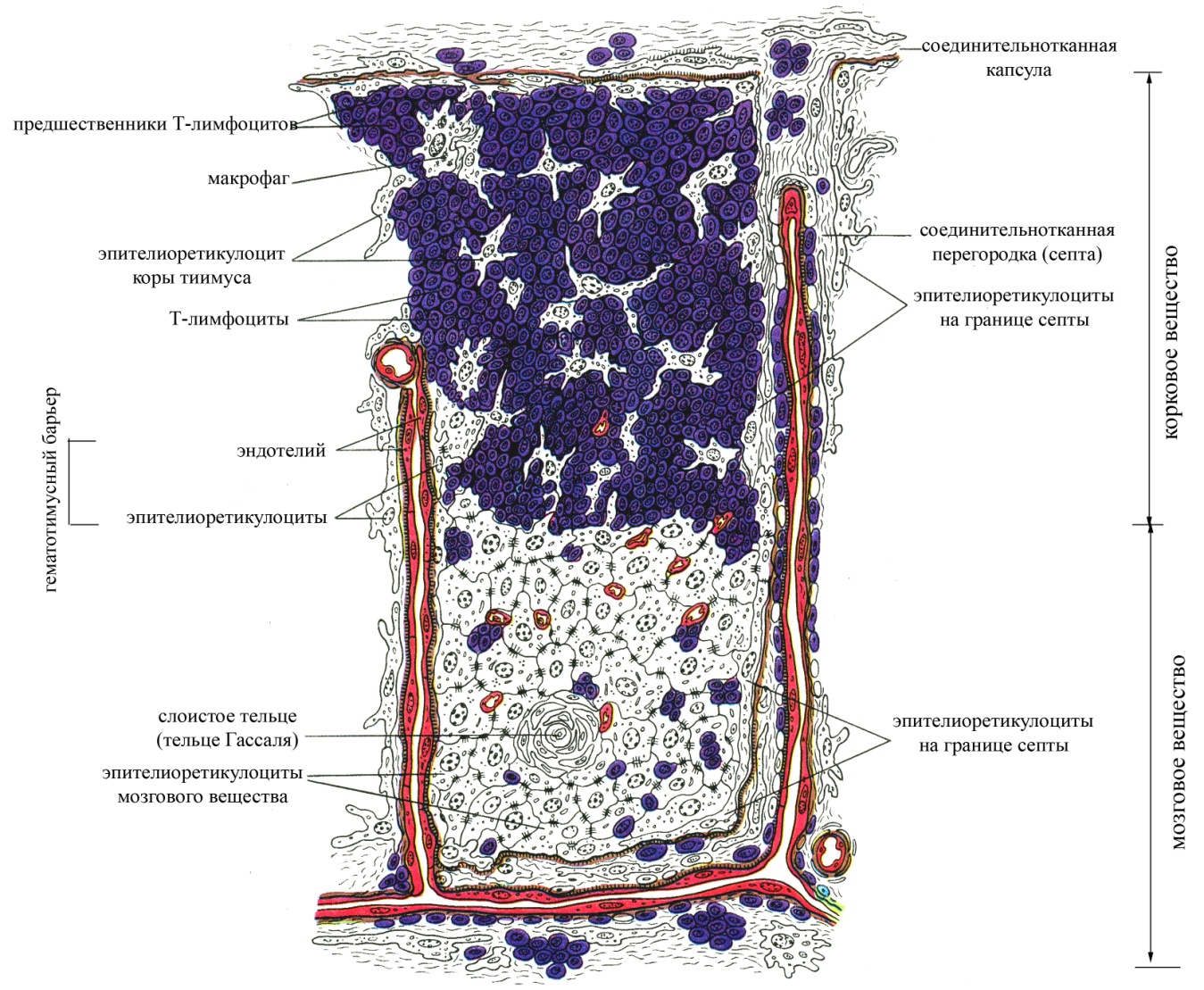
Рис. 10. Структурная организация дольки тимуса
В паренхиме тимусе различают:
корковое вещество, располагается непосредственно под капсулой органа, имеет более плотную консистенцию, образовано эпителиальными древовидными клетками, контактирующими отростками и формирующими таким образом сеть. В ячеях этой сети располагается большое количество лимфоцитов на разных стадиях дифференцировки. Причем самое периферическое положение в тимусе занимают наиболее крупные и наименее дифференцированные лимфоидные клетки – лимфобласты, для которых характерны частые митозы (эти клетки располагаются ближе всего к капсуле тимуса). По мере созревания размеры лимфоидных клеток уменьшаются, сокращается их митотическая активность, и они перемещаются ближе к центральной части тимуса (т.е. в направлении мозгового вещества). Таким образом, корковое вещество тимуса является автономной "фабрикой" Т-лимфоцитов, где особенно в эмбриональный и ранний постнатальный период осуществляется интенсивное их образование. Т-лимфоциты, образующиеся в корковом веществе тимуса, в дальнейшем заселяют другие лимфоидные органы, в которых превращаются в окончательно зрелые формы. Ежедневное количество новообразованных в корковом веществе Т-лимфоцитов, выходящих из тимуса, очень велико и приблизительно в 4 раза превышает нормальную общую популяцию лимфоцитов крови. Эпителиальные древовидные клетки в корковом веществе тимуса со всех сторон окружают развивающиеся лимфоидные клетки, принимая участие в создании здесь гематотимического барьера, образованного
эндотелиальными клетками капилляров с окружающей их базальной мембранной,
периваскулярной соединительной тканью, содержащей тканевую жидкость и макрофаги,
отростками эпителиальных древовидных клеток коркового вещества тимуса.
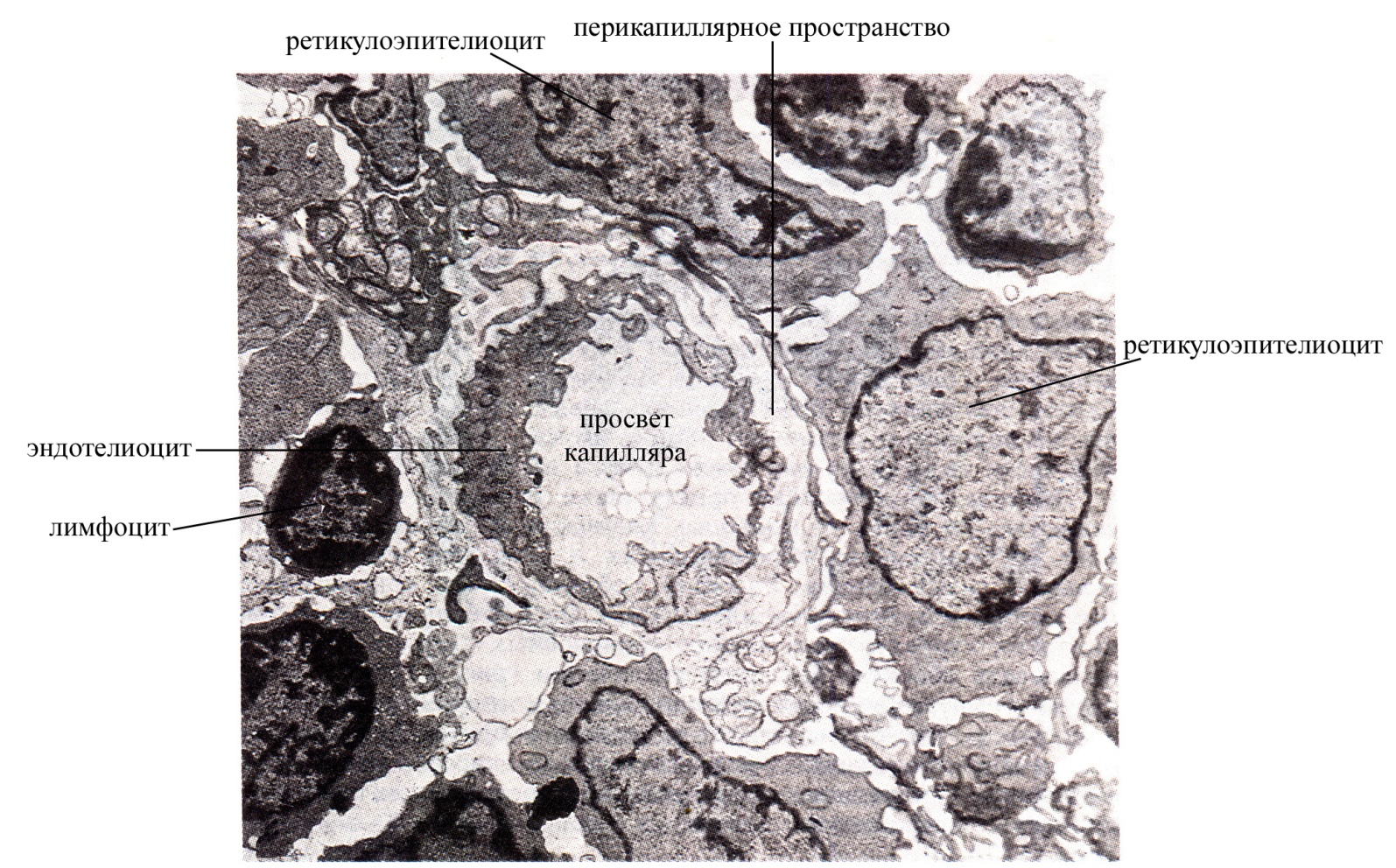
Рис. 11. Ультраструктура гематотимического барьера в корковом веществе тимуса (вид при электронной микроскопии)
Благодаря гематотимическому барьеру в корковом веществе тимуса исключается возможный контакт развивающихся лимфоцитов с антигенами внутренней среды организма, приносимыми кровью. Важным для развивающихся в корковом веществе Т-лимфоцитов является и тот факт, что в капсулу тимуса, в отличие от лимфатического узла не входят лимфатические сосуды, в результате чего лимфа не проходит через тимус, и это также исключает возможность доставки в корковое вещество органа чужеродных антигенов. Ранний контакт Т-лимфоцитов, коммитированных на какой-то антиген, с этим антигеном в корковом веществе тимуса сопровождается уничтожением такого клона Т-лимфоцитов. Благодаря уничтожению определенных клонов Т-лимфоцитов на уровне коркового вещества тимуса непосредственно после их возникновения и еще до момента их расселения в другие лимфоидные органы становится возможным выработка иммунологической толерантности по отношению к собственным антигенам гистосовместимости, широко представленным на поверхности эпителиальных древовидных клеток, окружающих развивающиеся лимфоидные клетки в корковом веществе. Вместе с тем наличие гематотимического барьера в корковом веществе тимуса, исключающее возможность контакта чужеродных антигенов, циркулирующих в крови, с развивающимися лимфоидными клетками, препятствует уничтожению клонов Т-лимфоцитов, коммитированных на чужеродные антигены и формированию иммунологической толерантности по отношению к чужеродным антигенам. В опытах на животных показано, что введение непосредственно в корковое вещество тимуса какого-то чужеродного антигена в эмбриональный период еще до момента расселения Т-лимфоцитов из тимуса в другие лимфоидные органы (селезенку и лимфатические узлы), сопровождается формированием иммунологической толерантности по отношению к этому антигену (т.е. такой антиген не воспринимается организмом как чужеродный и не вызывает иммунных реакций). Около 95% вновь образующихся в корковом веществе тимуса Т-лимфоцитов погибает, поскольку они вероятнее всего коммитированы на собственные антигены тканей организма, и лишь 5% лимфоцитов поступает в общий кровоток и далее оседает в лимфатических узлах и селезенке, где они могут контактировать с чужеродными антигенами. Часть зрелых Т-лимфоцитов из коркового вещества тимуса мигрирует в мозговое с током венозной крови, оттекающей от коркового вещества благодаря определенным особенностям кровоснабжения тимуса. В частности, артериолы тимуса проходят на границе коркового и мозгового вещества и дают начало самостоятельным капиллярным сетям этих двух зон тимуса. Вместе с тем венозная кровь, оттекающая от коркового вещества, собирается в венулы и вены мозгового, благодаря чему обязательно проходит через мозговое вещество тимуса. Кроме того, в мозговое вещество тимуса Т-лимфоциты могут приноситься с артериальной кровью, поступающей в орган (рециркулирующие полностью зрелые Т-лимфоциты). Каждый образующийся в корковом веществе тимуса Т-лимфоцит заранее коммитирован на какой-то определенный антиген (на плазматической мембране зрелых Т-лимфоцитов расположены рецепторы, которые могут избирательно взаимодействовать лишь с каким-то строго специфичным антигеном).
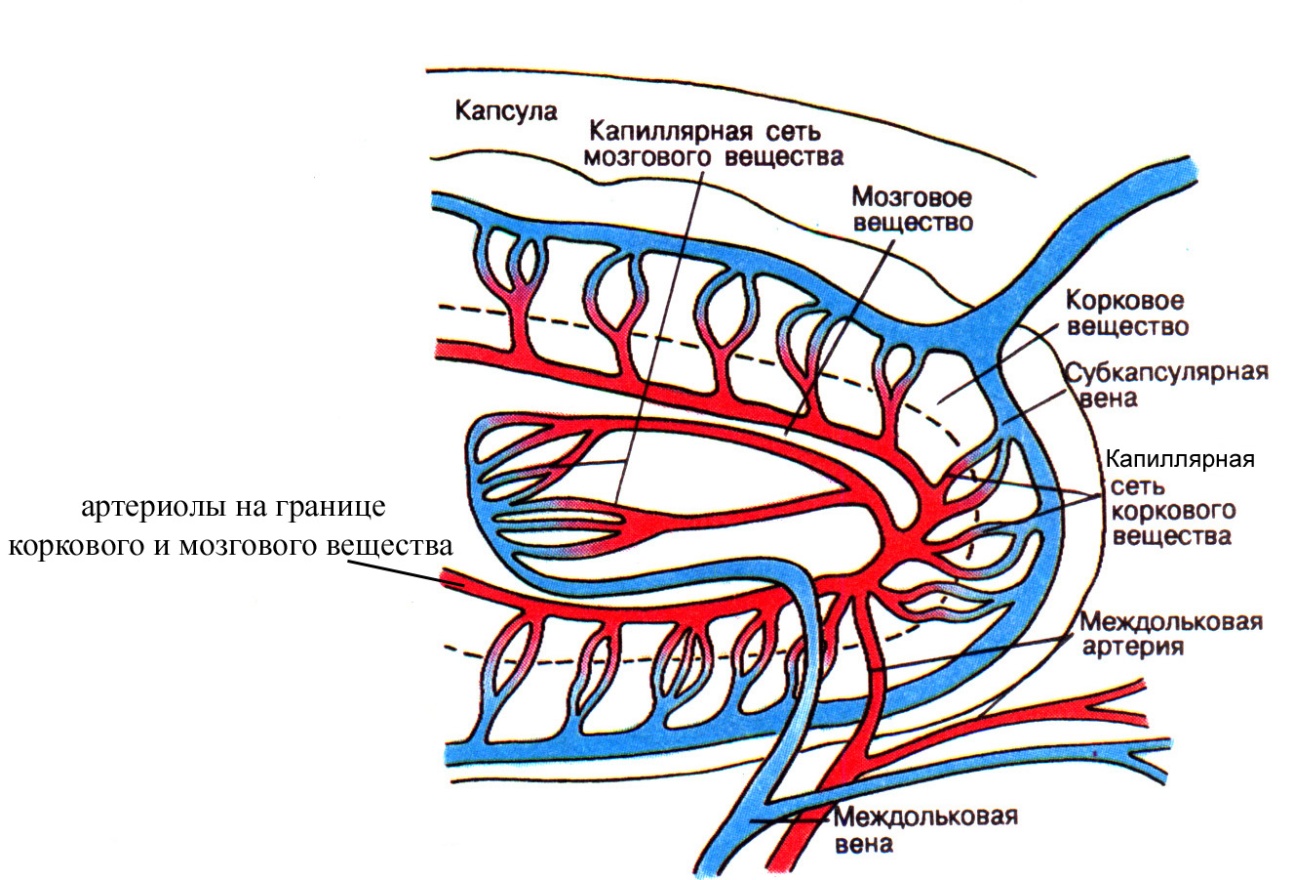
Рис. 12. Схема кровоснабжения дольки тимуса. Обратите внимание, что артериолы не проникают вглубь коркового вещества тимуса, а проходят, как правило, на границе коркового и мозгового вещества, разветвляясь на капиллярные сети коркового и мозгового вещества.
мозговое вещество, занимает центральное положение в тимусе, характеризуется более рыхлой консистенцией, образовано сетью древовидных эпителиальных клеток и расположенных между ними малых лимфоцитов (зрелых форм лимфоцитов). Для мозгового вещества тимуса характерна меньшая плотность лимфоцитов. Эпителиальная паренхима мозгового вещества тимуса представлена двумя компонентами:
диффузно расположенными древовидными эпителиоцитами, формирующими сеть, подобную таковой коркового вещества,
плотно упакованными эпителиоцитами, образующими слоистые тельца в мозговом веществе (называемые тельцами Гассаля), округлой формы, диаметром до 100 мкм; в таких тельцах многочисленные слои эпителиальных клеток располагаются вокруг одной центральной клетки тельца. Появляются эти тельца в тимусе на 12-ой неделе эмбрионального развития, их количество достигает максимума к 11 годам, а с 15 лет начинает уменьшаться. Роль этих телец до конца не ясна. Было замечено, что после введения в организм антигенов в тельцах происходят определенные циклические изменения и обнаруживается антиген. Существует предположение, согласно которому клетки, образующие тельца Гассаля, обладают фагоцитарной способностью.
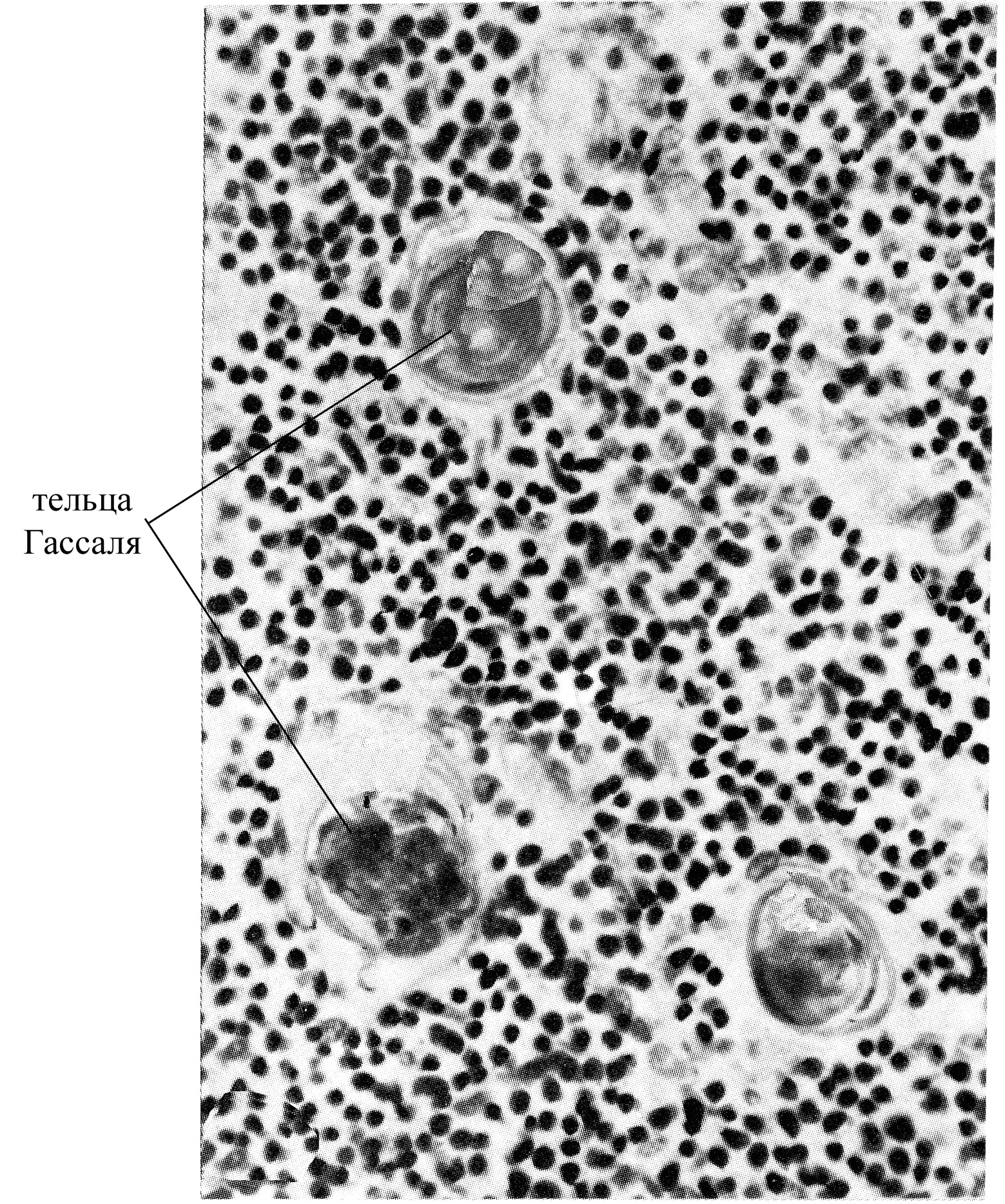
Рис. 13. Микрофотография участка мозгового вещества тимуса при большом увеличении (видны слоистые тельца или тельца Гассаля)
Эпителиальная паренхима тимуса, наряду с опорной функцией и созданием определенного микроокружения (представленного собственными антигенами гистосовместимости) вокруг Т-лимфоцитов, выполняет и эндокринную функцию. В частности, эпителиальные клетки тимуса синтезируют пептидные гормоны – тимозины и тимопоэтин. Функции тимозинов сводятся к следующему:
способствуют дифференцировке Т-лимфоцитов и появлению специфических рецепторов в их плазматической мембране (т.е. способствует развитию специфичности Т-лимфоцитов в отношении определенных антигенов),
стимулируют выработку многих цитотоксических гормонов лейкоцитами (т.н. лимфокинов, в том числе интерлейкина-2 Т-хелперами)
стимулируют продукцию иммуноглобулинов В-лимфоцитами и плазматическими клетками.
Другой гормон тимуса – тимопоэтин – является стимулятором дифференцировки предшественников Т-лимфоцитов, способствует дифференцировке самих Т-лимфоцитов, но в отличие от тимозинов, не влияет на развитие специфичности Т-лимфоцитов (т.е. на их коммитированность в отношении определенных антигенов).
Неинкапсулированные лимфатические фолликулы в рыхлой волокнистой соединительной ткани
В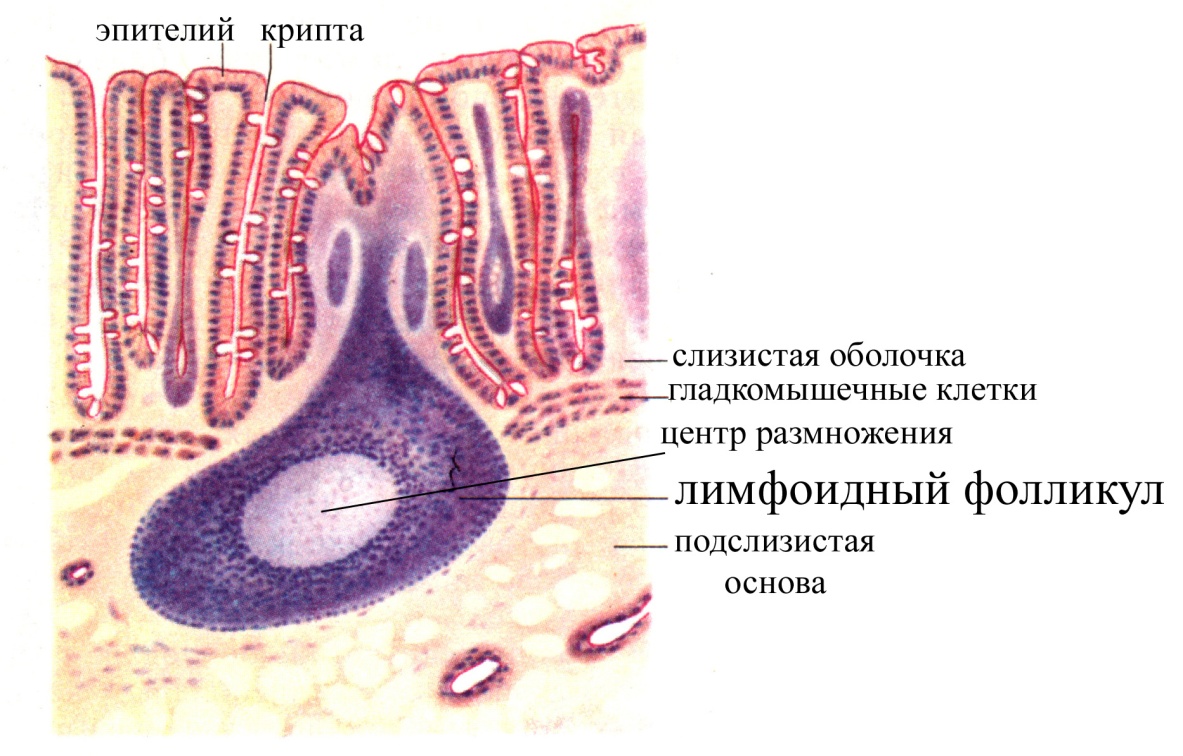 рыхлой волокнистой соединительной
ткани, расположенной под пограничным
эпителием (эпителием слизистой полых
органов – дыхательной, пищеварительной,
мочеполовой трубок и в других областях
организма) встречаются неинкапсулированные
лимфоидные фолликулы,
которые, как правило, находятся в тесной
связи с эпителиальной тканью (расположены
непосредственно под ней). Фолликулы
имеют округлую
форму,
диаметр – до 1 мм. Большинство лимфатических
фолликулов является дискретными, но в
некоторых областях организма они
образуют скопления, сливаясь в единую
массу. Так, в области языка, глотки и
носоглотки скопления лимфоидных
фолликулов формируют миндалины,
в рыхлой волокнистой соединительной
ткани кишечника – пэйеровы
бляшки.
Основными структурами лимфатических
фолликулов являются дифференцированные
малые лимфоциты и производные В-лимфоцитов
– плазматические клетки, которые
способны покидать фолликул с периферической
его области и заселять окружающую рыхлую
волокнистую соединительную ткань.
рыхлой волокнистой соединительной
ткани, расположенной под пограничным
эпителием (эпителием слизистой полых
органов – дыхательной, пищеварительной,
мочеполовой трубок и в других областях
организма) встречаются неинкапсулированные
лимфоидные фолликулы,
которые, как правило, находятся в тесной
связи с эпителиальной тканью (расположены
непосредственно под ней). Фолликулы
имеют округлую
форму,
диаметр – до 1 мм. Большинство лимфатических
фолликулов является дискретными, но в
некоторых областях организма они
образуют скопления, сливаясь в единую
массу. Так, в области языка, глотки и
носоглотки скопления лимфоидных
фолликулов формируют миндалины,
в рыхлой волокнистой соединительной
ткани кишечника – пэйеровы
бляшки.
Основными структурами лимфатических
фолликулов являются дифференцированные
малые лимфоциты и производные В-лимфоцитов
– плазматические клетки, которые
способны покидать фолликул с периферической
его области и заселять окружающую рыхлую
волокнистую соединительную ткань.
Рис. Лимфоидный фолликул слизистой тонкого кишечника
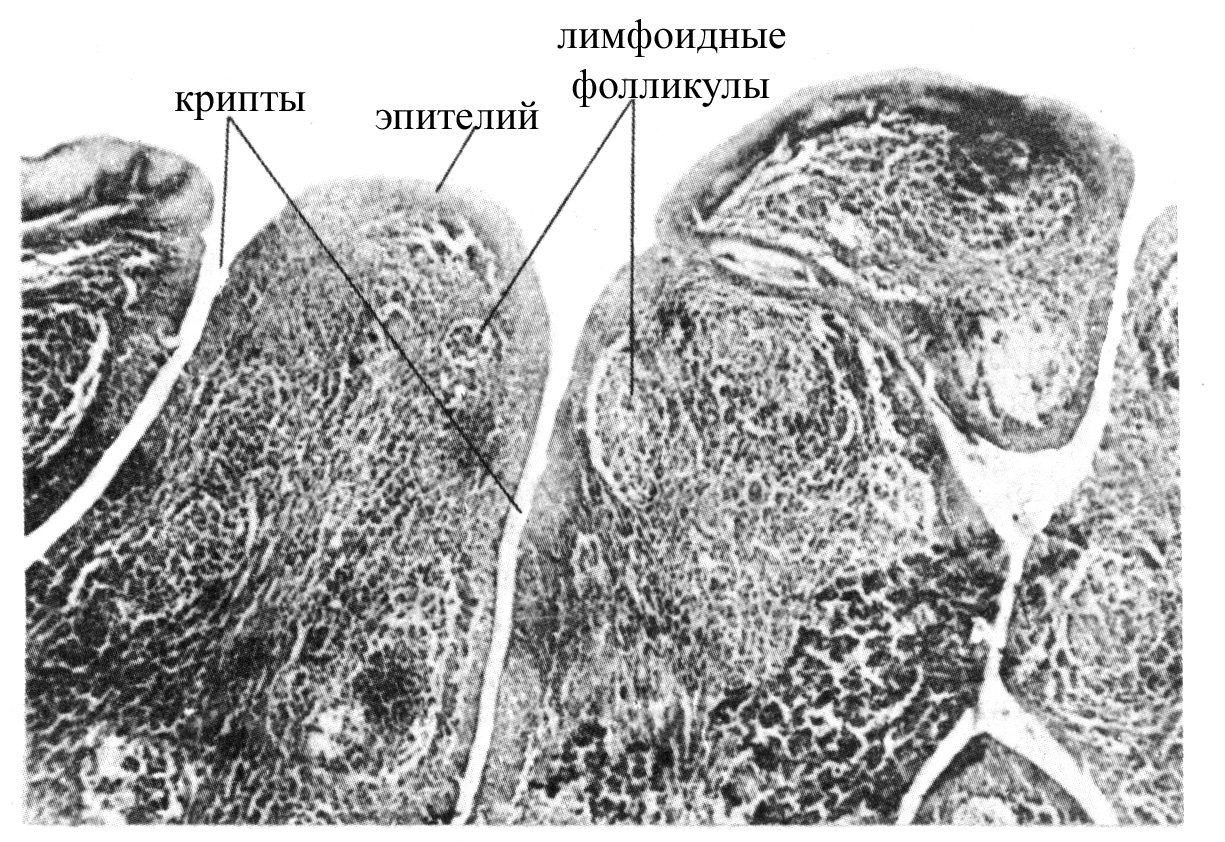
Рис. Скопление лимфоидных фолликулов в небных миндалинах человека
Лимфатические фолликулы начинают формироваться еще в эмбриональном периоде, но количество их резко увеличивается сразу после рождения и в последующей постнатальной жизни в результате встречи лимфоцитов с антигенами, проникающими под эпителиальный пласт в соединительную ткань. Следовательно, стимулом для формирования лимфатических фолликулов является контакт малых лимфоцитов (в основном В-лимфоцитов) с антигенами, в результате чего лимфоциты начинают пролиферировать, В-лимфоциты дают начало плазматическим клеткам. Пролиферация и дифференцировка В-лимфоцитов после антигенной стимуляции и являются причиной формирования лимфатического фолликула. В связи с отмеченным, у животных, выращенных в стерильных условиях (т.н. "безмикробных животных") количество лимфатических фолликулов очень мало.
Функциональное значение лимфатических фолликулов состоит в том, что они представляют собой второй (после пограничного эпителия) эшелон защиты организма от проникающих под эпителиальный пласт антигенов.
Лимфатические узлы
Лимфатические узлы являются периферическими органами иммунной системы, в которых осуществляется
контакт лимфоцитов с антигенами, сопровождающийся активацией лимфоцитов. Процесс активации лимфоцитов, в свою очередь, включает следующие реакции:
антигензависимую пролиферацию (клонирование) определенных лимфоцитов, детерминированных на данный антиген,
последующую дифференцировку Т- и В-лимфоцитов в эффекторные клетки. В частности, В-лимфоциты превращаются в плазматические клетки, а Т-лимфоциты – в Т-хелперы или Т-киллеры (цитотоксические лимфоциты)
образование клеток-памяти (Т- или В-клеток памяти в результате активации лимфоцитов после контакта с определенным антигеном)
очистка лимфы от инородных частиц и антигенов (на 95-99%), связанная с активацией лимфоцитов и присутствием макрофагов, а также освобождение лимфы от избытка воды, белков, жиров, обогащение ее антителами и лимфоцитами
депонирование лимфы и некоторые другие процессы.
Следовательно, лимфатические узлы служат местом лимфоцитопоэза (причем не первичного (антигеннезависимого) из кроветворных стволовых клеток, которое осуществляется в центральных органах иммунной системы (красном костном мозге и тимусе), а вторичного антигензависимого образования активированных Т- и В-лимфоцитов из малодифференцированных лимфоцитов-предшественников, коммитированных на данный антиген), иммунной защиты и депонирования лимфы.
Лимфатические узлы имеют овально-округлую форму, диаметр – 0,5-1 см. Они располагаются по ходу крупных лимфатических сосудов, образуя большие скопления в подкрыльцовой впадине, паху, вдоль крупных сосудов шеи, в грудной клетке и брюшной полости, особенно по ходу крупных сосудов, в брыжейке и некоторых других областях организма. Следовательно, лимфатические узлы, в отличие от лимфатических фолликулов, локализованы не там, где образуется лимфа, а вдоль главных ветвей грудного и правого лимфатического протоков, через посредство которых лимфа возвращается в кровь. Обычно у лимфатических узлов с одной стороны имеется вдавление, называемое воротами. В области ворот в узел входят артерии и нервы, выходят вены и выносящие (эфферентные) лимфатические сосуды. Противоположная вдавлению сторона узла имеет выпуклую форму и называется наружной его стороной; она соответствует месту проникновения в лимфатический узел приносящих (аффрентных) лимфатических сосудов. Таким образом, лимфа входит в лимфатический узел со стороны наружной его поверхности, проходит через него и выходит по выносящим лимфатическим сосудам в области ворот узла.
Лимфатический узел покрыт соединительнотканной капсулой, содержащей большое количество жировых клеток. От капсулы узла со стороны его ворот отходят, направленные внутрь многочисленные соединительнотканные трабекулы, достигающие наружной поверхности узла, образующие в совокупности соединительнотканнную его строму и выполняющие опорную и трофическую функции.
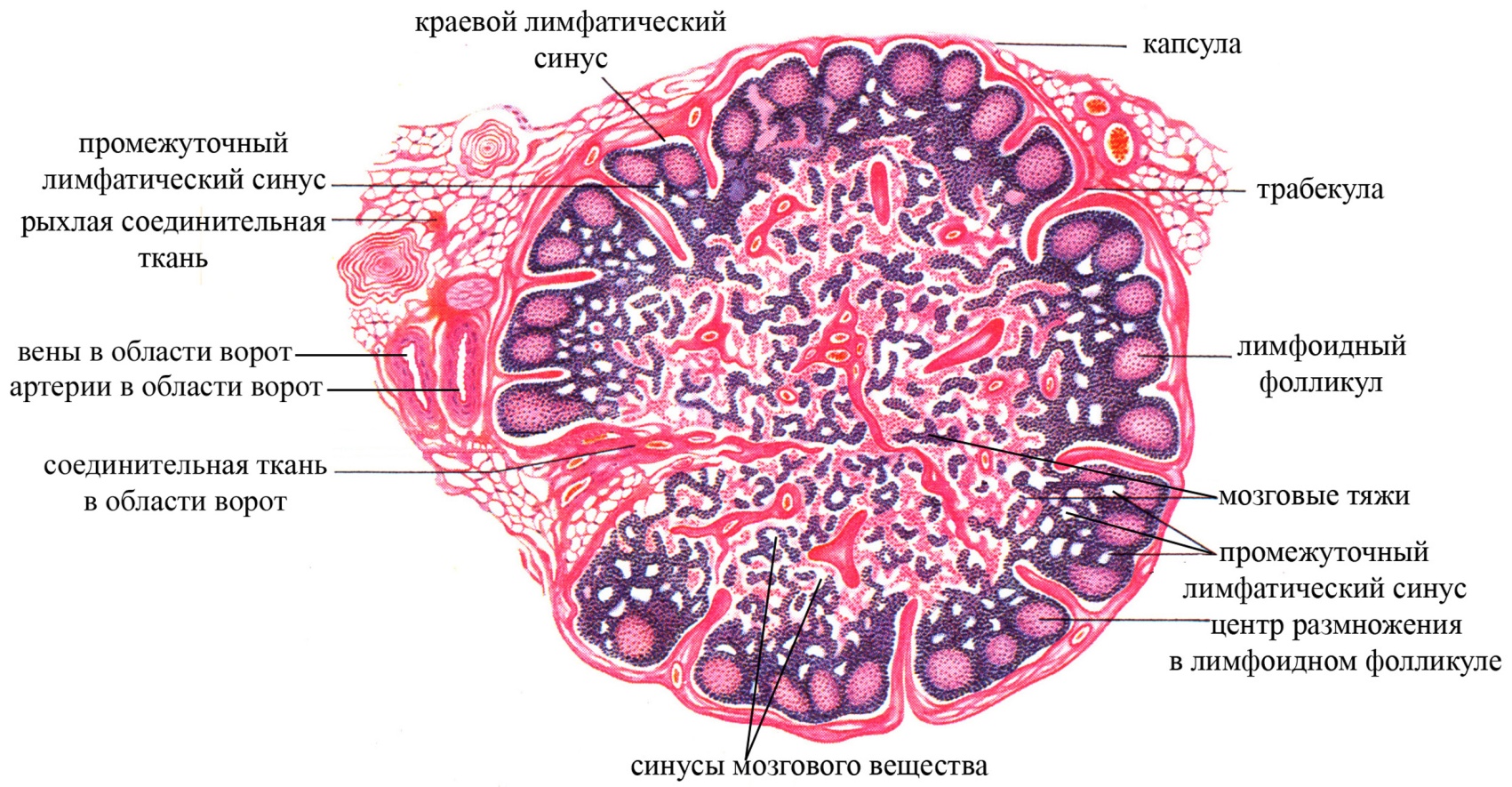
Рис. Гистоструктура лимфатического узла
В лимфатическом узле выделяют корковое и мозговое вещество, а также лимфатические синусы (полости в лимфатическом узле, определенным образом в нем локализующиеся, выстланные лимфатическим эндотелием или эндотелиоподобными ретикулярными клетками, заполнены лимфой).
К орковое
вещество
занимает
периферическое положение в узле, имеет
более плотную структуру, в периферической
своей части содержит лимфатические
фолликулы
(называемые также первичными лимфатическими
фолликулами и идентичные неинкапсулированным
лимфатическим фолликулам рыхлой
волокнистой соединительной ткани).
Лимфатические фолликулы имеют округлую
форму, диаметр до 0,5 мм, представлены
лимфоидными клетками на разных этапах
дифференцировки (преимущественно
В-лимфоцитами, среди которых в гораздо
меньшем количестве находятся Т-хелперы)
и макрофагами, залегающими в ретикулярной
ткани. Снаружи лимфатические фолликулы
покрыты эндотелиоподобными ретикулярными
клетками, лежащими на пучках ретикулярных
фибрилл. В толще коркового вещества в
радиальном направлении проходят
соединительнотканные трабекулы (являются
производными соединительнотканной
капсулы узла) и лимфатические синусы,
разделяя ретикулолимфоидную его
структуру на отдельные дольки, имеющие
форму пирамид (основание этих пирамид
обращено к соединительнотканной капсуле,
а вершины – к мозговому веществу и
переходят в мякотные тяжи). Именно в
пирамидах коркового вещества и залегают
лимфатические фолликулы.
орковое
вещество
занимает
периферическое положение в узле, имеет
более плотную структуру, в периферической
своей части содержит лимфатические
фолликулы
(называемые также первичными лимфатическими
фолликулами и идентичные неинкапсулированным
лимфатическим фолликулам рыхлой
волокнистой соединительной ткани).
Лимфатические фолликулы имеют округлую
форму, диаметр до 0,5 мм, представлены
лимфоидными клетками на разных этапах
дифференцировки (преимущественно
В-лимфоцитами, среди которых в гораздо
меньшем количестве находятся Т-хелперы)
и макрофагами, залегающими в ретикулярной
ткани. Снаружи лимфатические фолликулы
покрыты эндотелиоподобными ретикулярными
клетками, лежащими на пучках ретикулярных
фибрилл. В толще коркового вещества в
радиальном направлении проходят
соединительнотканные трабекулы (являются
производными соединительнотканной
капсулы узла) и лимфатические синусы,
разделяя ретикулолимфоидную его
структуру на отдельные дольки, имеющие
форму пирамид (основание этих пирамид
обращено к соединительнотканной капсуле,
а вершины – к мозговому веществу и
переходят в мякотные тяжи). Именно в
пирамидах коркового вещества и залегают
лимфатические фолликулы.
Рис. Гистоструктура коркового вещества лимфатического узла
Мозговое вещество занимает центральное положение в лимфатическом узле, представлено мякотными тяжами, которые являются продолжением коркового вещества, имеет более рыхлую структуру по сравнению с коровым веществом.
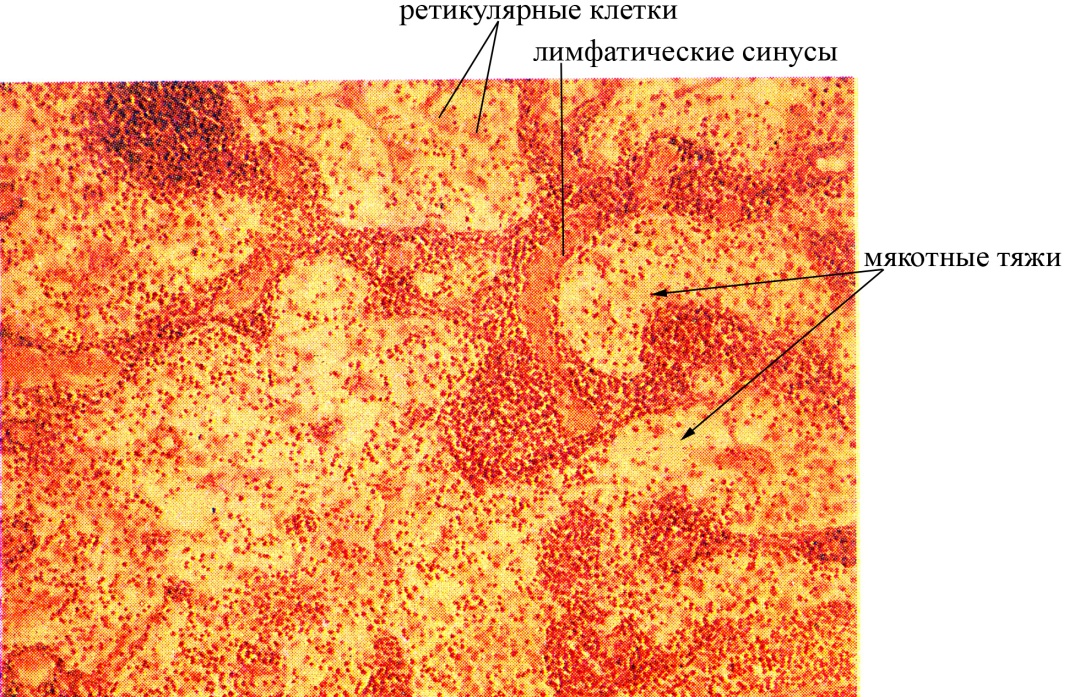
Рис. Гистоструктура мозгового вещества лимфатического узла
В основе коркового и мозгового вещества лимфатических узлов лежит ретикулярная ткань и лимфоидные клетки, находящиеся на разных этапах дифференцировки. Кроме того, и в мозговом, и в корковом веществе лимфатических узлов содержится большое количество макрофагов. Ретикулярная ткань лимфатического узла представлена дендритными ретикулярными клетками и ретикулиновыми волокнами, контактирующими между собой и образующими подобие петлистой сети, в ячеях которой локализованы лимфоидные клетки. Ретикулиновые волокна соединяются не только между собой, но и с коллагеновыми и эластическими волокнами соединительной ткани капсулы узла и его трабекул, что обеспечивает создание прочного волокнистого каркаса внутри лимфатического узла. Лимфоидные клетки коркового и мозгового вещества узла оказываются окруженными со всех сторон дендритными ретикулярными клетками, которые выполняют следующие функции:
антигенпредставляющую (связывают антигены и локализуют их на своей поверхности; благодаря большой суммарной площади своей поверхности (за счет наличия большого количества отростков) они обеспечивают поддержание высоких концентраций антигенов вокруг созревающих клеток лимфоидного ряда)
трофическую по отношению к клеткам лимфоидного ряда (принимают участие в доставке различных веществ из крови к лимфоидным клеткам и в обратном направлении), в связи с чем их образно называют "клетками-няньками" лимфоидных клеток
опорную по отношению к развивающимся лимфоидным клеткам.
Наряду с ретикулярными клетками лимфоидные клетки коркового и мозгового вещества лимфатического узла окружены большим количеством макрофагов, часть из которых обладает способностью к фагоцитозу, а часть – утратила эту способность. Макрофаги, участвующие в фагоцитозе, характеризуются наличием в своей цитоплазме большого количества фагосом, они, с одной стороны, способствуют очистке протекающей через узел лимфы от различных антигенов, а с другой – принимают участие в иммунном ответе, поскольку обеспечивают превращение корпускулярного антигена в молекулярный, который доступен для различения иммунокомпетентными лимфоцитами. Так, макрофаги после фагоцитоза антигенов частично их лизируют, после чего транспортируют на свою поверхность какие-то молекулярные компоненты этих антигенов, где они связываются с рецепторами мембраны макрофага и могут различаться определенными иммунокомпетентными лимфоцитами, коммитированными на данный антиген. Часть макрофагов, находящихся в лимфатическом узле утратила способность к фагоцитозу. Существует предположение, согласно которому такие клетки узла являются "пришлыми" макрофагами, потомками клеток Лангерганса кожи, мигрирующими в узел и приносящими сюда антигены, поступившие в организм через поверхность кожи. Причем эти макрофаги характеризуются большой продолжительностью жизни и специализируются на антигенпредставляющей функции, поскольку фиксируют на своей поверхности различные молекулярные компоненты антигенов, в том числе проникшие через кожу.
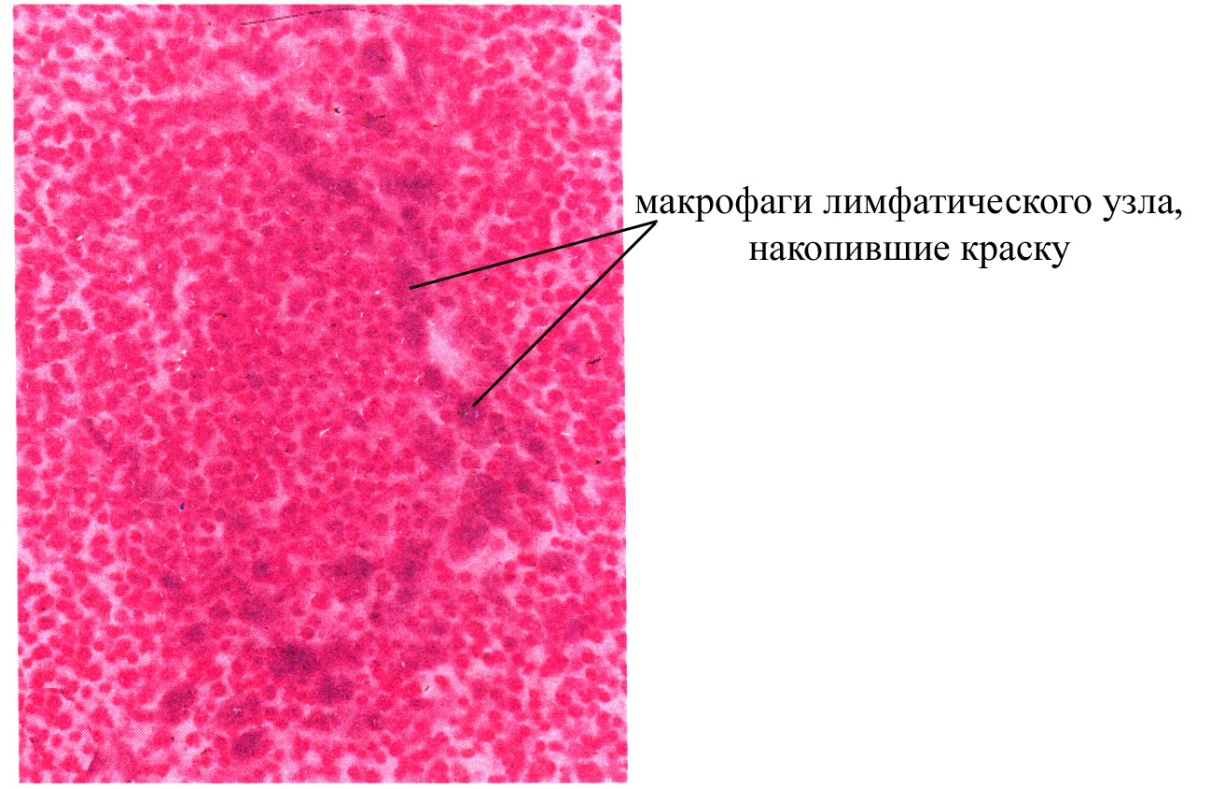
Рис. Накопление краски в макрофагах лимфатического узла (окраска гематоксилин-эозином)
Распределение популяций Т- и В-лимфоцитов в корковом и мозговом веществе лимфатического узла подчиняется определенной закономерности. Так, в периферической части коркового вещества, соответствующей месту локализации лимфатических фолликулов, преобладают в основном В-лимфоциты, а сами эти лимфатические фолликулы являются местом антигензависимой активации В-лимфоцитов, в которой участвуют Т-хелперы и образуются активированные В-лимфоциты. Часть активированных В-лимфоцитов мигрирует в мозговое вещество, где на вершинах мякотных тяжей превращается в плазматические клетки, продуцирующие антитела, а часть – дифференцируется в В-клетки памяти, которые могут оставаться в узле или поступать в кровь и лимфу и мигрировать по организму. Плазматические же клетки, в отличие от В-клеток памяти являются неподвижными и так и остаются в области мякотных тяжей, продуцируя антитела.
Следовательно, лимфатические фолликулы являются местом антигензависимого дифференцирования В-лимфоцитов, которое протекает с участием макрофагов (антигенпредставляющих клеток) и в большинстве случаев – Т-хелперов. Состояние этих фолликулов во многом зависит от проникновения определенных антигенов в узел. В частности, после антигенной стимуляции в некоторых лимфатических фолликулах появляются более светлые зоны, называемые центрами размножения. Они соответствуют месту нахождения крупных лимфоидных клеток, активно делящихся и дающих начало активированным малым лимфоцитам. Поскольку в центре узла содержатся преимущественно крупные предшественники малых лимфоцитов, эта область узла имеет более светлую окраску, обусловленную относительно большим объемом цитоплазмы крупных лимфоидных клеток, по сравнению с активированными малыми лимфоцитами. Периферия же лимфоидных фолликулов представлена скоплениями преимущественно активированных малых лимфоцитов, характеризующихся меньшими размерами и объемом цитоплазмы и поэтому создающих более темную картину в связи с большей плотностью ядер. В отсутствии антигенов в узле большая часть первичных фолликулов не имеет центров размножения. Эти центры отсутствуют также до момента рождения и у "безмикробных" животных. Таким образом, лимфатический узел представляет собой очень динамичную структуру, в которой, в зависимости о проникающих антигенов, происходит возникновение центров размножения в определенных лимфатических фолликулах на фоне временного "молчания" других.
Наряду с заселением В-лимфоцитами периферической части коркового вещества лимфатического узла, многочисленные их популяции встречаются и в мозговом веществе (среди ретикулярной ткани мякотных тяжей). В-лимфоциты мозгового вещества в большинстве своем являются активированными в результате контакта с определенными антигенами малыми лимфоцитами, мигрирующими из коркового вещества узла и находящимися на завершающих стадиях созревания. Они могут превращаться либо в В-лимфоциты памяти, либо в плазматические клетки (как правило, на вершине мякотных тяжей), продуцирующие в основном иммуноглобулины класса G. Наряду с В-лимфоцитами в ячеях ретикулярной ткани мозгового вещества содержаться и макрофаги. Внутри мякотных тяжей коркового вещества проходят артериолы и капилляры, содержащие поры в эндотелии, через посредство которых активированные В-лимфоциты могут поступать в кровоток. Снаружи мякотные тяжи, подобно лимфатическим фолликулам покрыты эндотелиоподобными ретикулярными клетками, лежащими на ретикулярных волокнах.
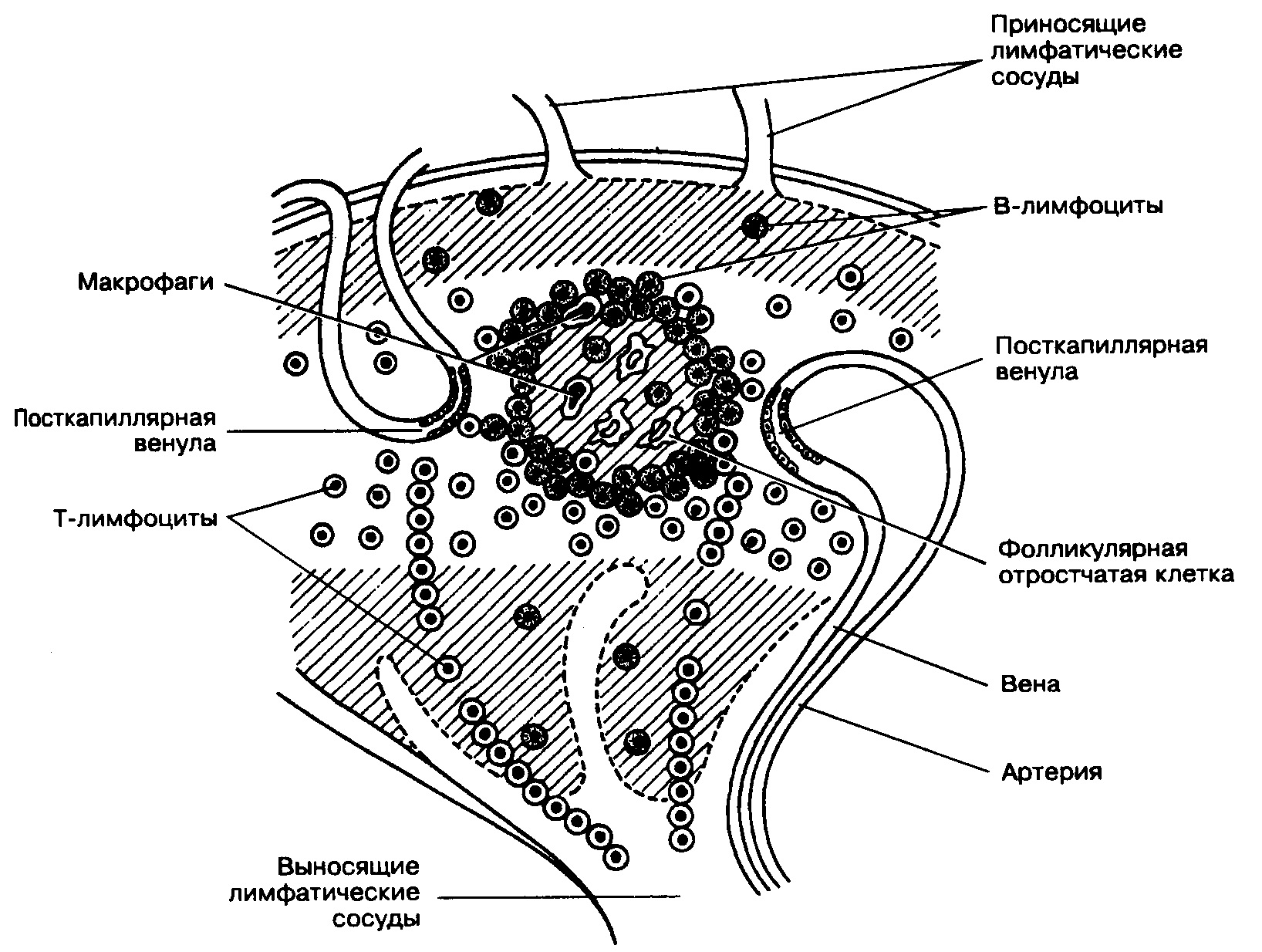
Рис. Схема распределения Т- и В-лимфоцитов в лимфатическом узле
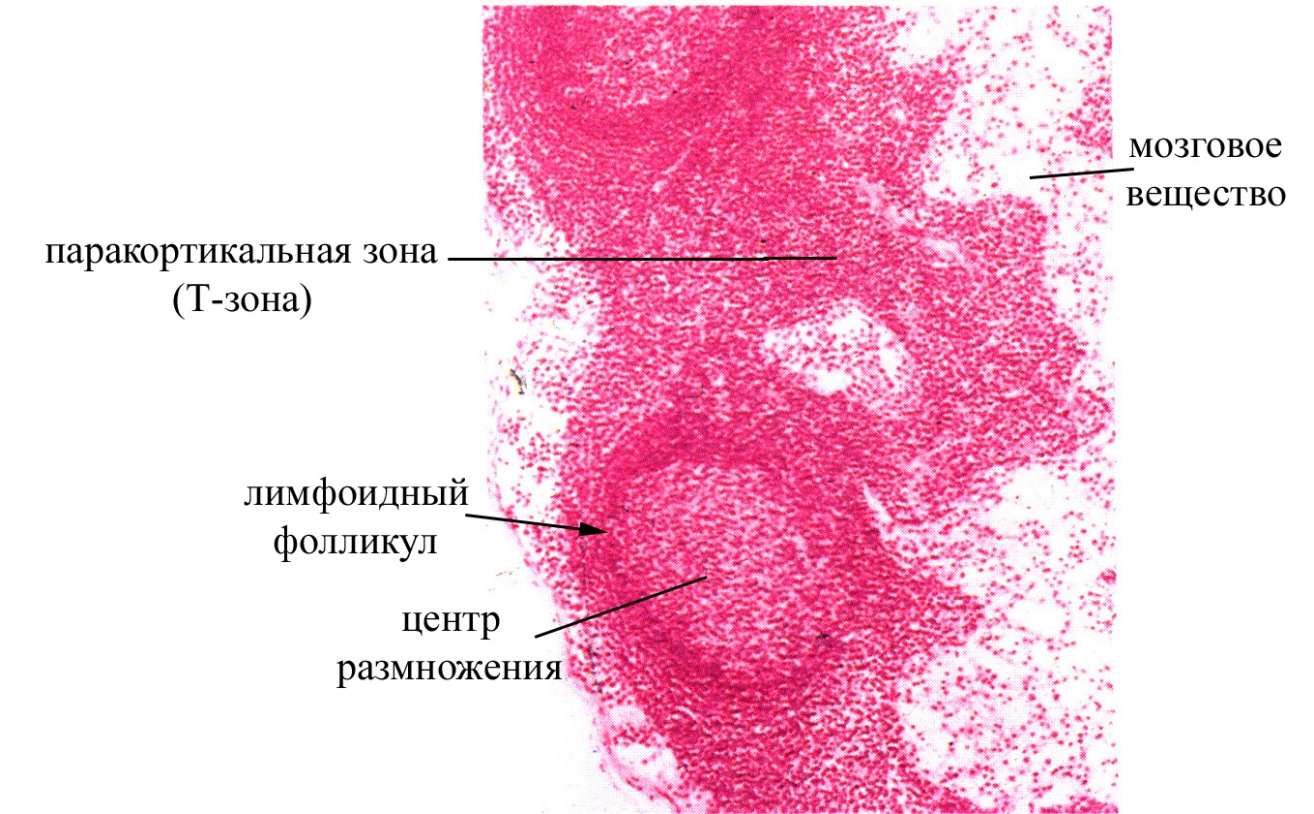
Рис. Локализация паракортикальной (Т-зоны) в лимфатическом узле
Наконец, зона узла, соответствующая средней и внутренней части коркового вещества и граничащая с мозговым веществом, называется тимусзависимой зоной узла, поскольку заселена преимущественно Т-лимфоцитами. Иногда эту зону узла называют еще паракортикальной, считая в таком случае истинной корой узла периферическую его часть, содержащую лимфатические фолликулы (т.е. тимусзависимая зона с одной своей стороны прилежит к периферической части коркового вещества узла, а с другой – контактирует с мозговым его веществом). В этой зоне узла в ячеях ретикулярной ткани преобладают Т-лимфоциты-хелперы, и несколько реже встречаются другие популяции Т-лимфоцитов (в частности, Т-киллеры). Тимусзависимая зона узла получила свое название в связи с тем, что заселяется она малодифференцированными Т-лимфоцитами, приносящимися из тимуса, и после удаления тимуса она запустевает из-за убыли Т-лимфоцитов. В этой зоне происходит антигензависимая пролиферация Т-клеток и дифференцировка в определенные эффекторные клетки (Т-киллеры, Т-хелперы и т.д.). Пересадка аллотрансплантанта вблизи места расположения лимфатического узла сопровождается резким увеличением тимусзависимой зоны, в связи с поступлением в узел чужеродных антигенов аллотрансплантанта и вызванной этим антигензависимой пролиферацией и активацией малодифференцированных Т-лимфоцитов, поступивших из тимуса и коммитированных на данные антигены. Наряду с ретикулярными и лимфоидными клетками в тимусзависимой зоне узла присутствуют и макрофаги, в том числе утратившие способность к фагоцитозу (т.н. интердигитирующие клетки). Интердигитирующие клетки обладают многочисленными пальцевидными отростками, вдавливающимися из одной клетки в другую, они мигрируют с током лимфы из кожи и являются производными внутриэпидермальных макрофагов (клеток Лангерганса), доставляющих в лимфатический узел антигены, проникшие через кожу. Функциональное значение таких интердигитирующих клеток состоит в том, что они специализируются на синтезе гликопротеидов примембранных слоев (т.е. гликопротеидов гликокаликса), которые способны сорбировать и сохранять антигены на плазматических мембранах, а также индуцировать пролиферацию Т-лимфоцитов.
Таким образом, распределение Т- и В-лимфоцитов в лимфатических узлах подчиняется следующей закономерности:
В-лимфоциты располагаются преимущественно в наружной части коркового вещества лимфатического узла и в области мякотных тяжей мозгового вещества. Причем В-лимфоциты коркового вещества в большинстве своем представляют собой малодифференцированные лимфоидные клетки, находящиеся на разных стадиях антигензависмой дифференцировки. Тогда как тяжи узла заселены преимущественно зрелыми малыми В-лимфоцитами, заканчивающими свою дифференцировку и превращающимися либо в плазматические клетки (неподвижные клетки, продуцирующие антитела), либо в В-лимфоциты памяти
Т-лимфоциты преобладают в средней и внутренней области коркового вещества узла и представляют собой лимфоидные клетки, находящиеся на разных этапах антигензависимой активации.
Наряду с соединительнотканной стромой, ретикулярной и лимфоидной тканями, типичной морфологической особенностью лимфатического узла являются синусы (в переводе с латинского означает полое пространство), заполненные лимфой (лимфатические синусы). В лимфатическом узле различают следующие синусы:
краевой синус, располагается непосредственно под соединительнотканной капсулой узла, выстлан лимфатическим эндотелием (уплощенные эндотелиальные клетки), не имеющим базальной мембраны, но контактирующим в некоторых местах с коллагеновыми и ретикулиновыми волокнами. Причем со стороны соединительнотканной капсулы эндотелиальная выстилка синуса является сплошной, тогда как со стороны лимфатических фолликулов корокового вещества – прерывистой. В краевой синус лимфатического узла поступает лимфа, приносимая по афферентным лимфатическим сосудам, несущим лимфу из тканей и проникающим в узел со стороны наружной его поверхности (поверхности, противоположной воротам узла). Краевой синус содержит много свободных клеток, преимущественно макрофагов и лимфоцитов, которые могут проникать сюда из подлежащего коркового вещества лимфатического узла. Благодаря многочисленным макрофагам и лимфоцитам краевой синус обеспечивает фагоцитоз различных антигенов, поступающих с лимфой, а, следовательно, принимает участие в начальной фильтрации и очистке лимфы.
корковые и промежуточные корковые синусы, отходящие радиально от краевого синуса и проходящие в толще коркового вещества между ретикулярно-лимфоидными его компонентами, имеющими форму пирамид. В этих синусах в протекающую лимфу попадает еще некоторое количество лимфоцитов из коркового вещества узла
мозговые синусы, являются продолжением синусов коркового вещества в мозговом веществе узла; проходят между мякотными тяжами, по этим синусам лимфа направляется к выносящим лимфатическим сосудам, покидающим узел в области его ворот.
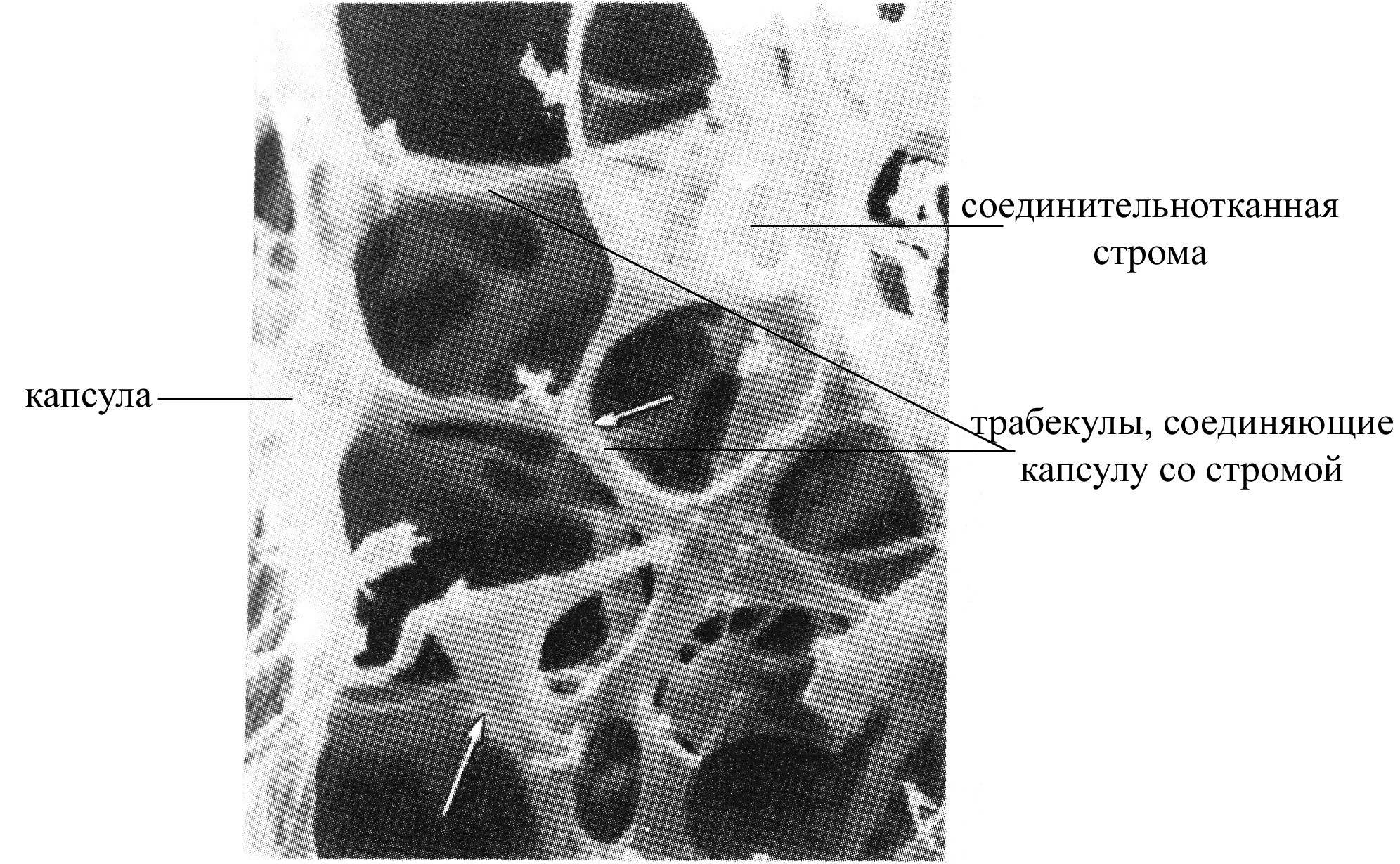
Рис. Краевой синус лимфатического узла (вид при сканирующей электронной микроскопии)
Стенка всех синусов устроена по общему принципу: она образована лимфатическим эндотелием или ретикулярными эндотелиоподобными клетками, не имеет базальной мембраны, укреплена в некоторых местах ретикулиновыми волокнами и характеризуется наличием крупных просветов между своими клетками, позволяющими клетками проходить в полость синуса или в обратном направлении в ткань узла. Обычными типами клеток внутри синусов являются лимфоциты и макрофаги, обеспечивающие очистку лимфы.
Кровоснабжение лимфатического узла осуществляется артериями, входящими в узел в области ворот. Эти артерии и более мелкие отходящие от них сосуды (мелкие артерии, артериолы) проходят в толще узла в составе соединительнотканных трабекул и мякотных тяжей и достигают коркового вещества узла. Причем артериолы до уровня лимфатических фолликулов в корковом веществе, как правило, не доходят, а разветвляются на капиллярную сеть на границе коркового и мозгового вещества. Таким образом, в периферической части коркового вещества лимфатического узла кровоснабжение обеспечивают в основном капилляры, которые переходят в посткапиллярные венулы. Посткапиллярные венулы следуют в корковом веществе в противоположном по отношению к капиллярам направлении (а именно в направлении из коркового вещества в мозговое, т.е. кровеносная сеть совершает обратный ход и возвращается в мозговое вещество). Посткапиллярные венулы переходят в мозговое вещество узла, где впадают в более крупные вены. Венозная кровь из лимфатического узла выносится, как правило, по общей его вене, покидающей узел в области ворот. Важной особенностью посткапиллярных венул коркового вещества лимфатического узла является относительно большая высота эндотелиальных клеток (гораздо большая по сравнению со многими другими венулами организма), в связи с чем такой эндотелий называют кубическим и даже высоким призматическим, а также наличие большого количества пор между эндотелиальными клетками. Функциональное значение такого высокого призматического эндотелия посткапиллярных венул состоит в том, что его клетки, после того как лимфоцит проник между ними, со стороны просвета сосуда (выше места его нахождения лимфоцита в стенке венулы) смыкаются и препятствует просачиванию жидкости. Таким образом, высокий призматический эндотелий венул осуществляет хоминг лимфоцитов, препятствуя при этом выходу плазмы из венулы в узел.
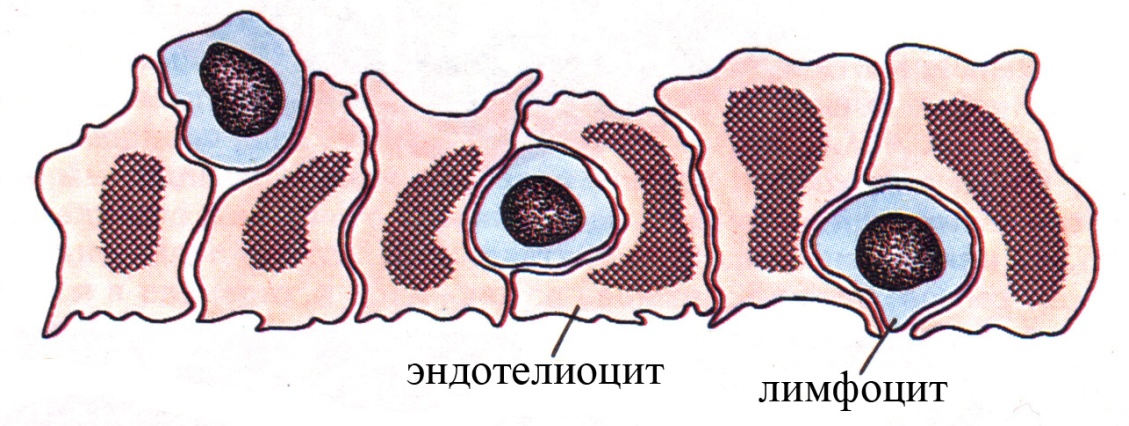
Рис. Миграция лимфоцитов сквозь стенку посткапиллярной венулы (схема)
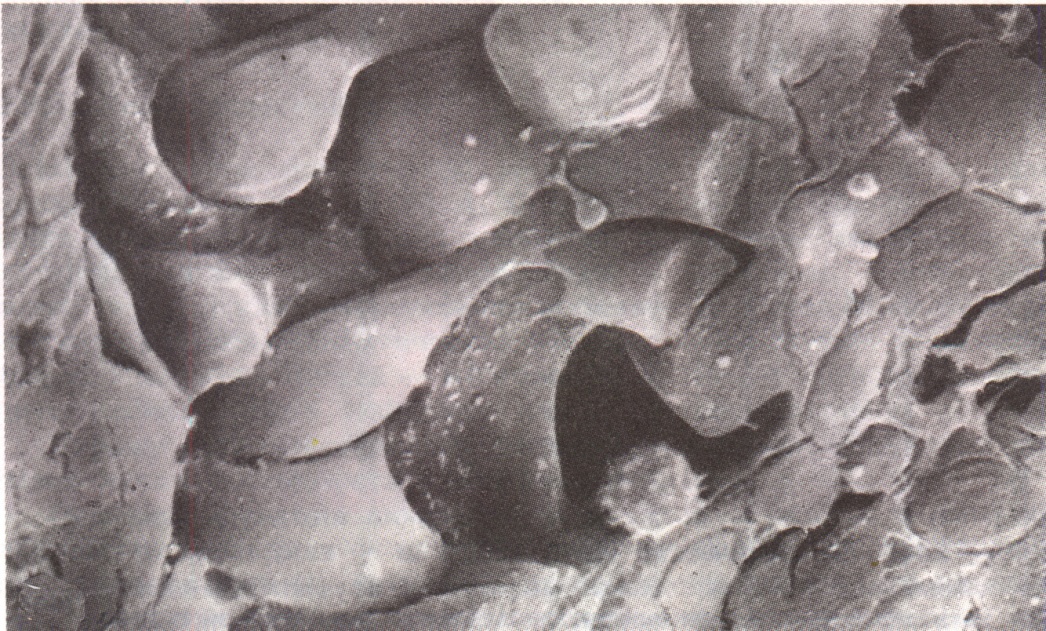
Рис. Миграция лимфоцитов сквозь стенку посткапиллярной венулы (сканирующая электронная микроскопия)
Лимфоциты, покинувшие посткапиллярные венулы, в большинстве своем поступают в лимфатические синусы или лимфоретикулярную ткань коркового и мозгового вещества, где могут контактировать с различными антигенами, принесенными лимфой из тканей, и в результате такого контакта активироваться. Кроме того, часть, вышедших из посткапиллярных венул лимфоцитов с током лимфы выносятся из узла и возвращаются обратно в кровь (рециркуляция лимфоцитов).
Развитие лимфатических узлов в эмбриональный период. Лимфатические узлы возникают в конце 2-го – начале 3-го месяца внутриутробного развития из мезенхимных клеток, образующих скопления в определенных областях кровеносных и лимфатических сосудов. Мезенхимные клетки дают начало соединительнотканной капсуле узла и трабекулам, а также ретикулярной ткани. К 16-й неделе зачатки лимфатических узлов начинают заселяться лимфоцитами. Причем В-лимфоциты вселяются раньше Т-лимфоцитов, главным образом в центральные участки лимфатического узла (будущее мозговое вещество), а затем в самый поверхностный (в дальнейшем – корковое вещество) слой. Т-лимфоциты вселяются в промежуточную зону между корковым и мозговым веществом (паракортикальную или тимусзависимую зону). На 16-й неделе в лимфатическом узле образуются лимфоидные фолликулы и мозговые тяжи, а также появляются ретикулярные волокна. к концу эмбриогенеза в лимфатических узлах заканчивается формирование всех структур – коркового вещества с лимфатическими фолликулами, мозговых тяжей, лимфатических синусов, Т- и В-зон. Однако до момента рождения (и соответственно начала контакта лимфоцитов узла с антигенами в его лимфатических фолликулах отсутствуют центры размножения, а плотность Т-зоны оказывается гораздо меньше, чем в постнатальном периоде).
Морфо-функциональная характеристика селезенки
Селезенка (splen, lien) представляет собой неполый орган иммунной системы, находящийся на пути крови из магистрального сосуда большого круга кровообращения – аорты – к печени (селезеночные вены впадают в воротную вену печени, вследствие чего, венозная кровь, вышедшая из селезенки, прежде чем попасть в нижнюю полую вену, проходит через печень). Она располагается в брюшной полости, внутрибрюшинно (покрыта висцеральным листком брюшины, являющейся серозной оболочкой селезенки), в области левого подреберья, на уровне от IХ до ХI ребер, занимает горизонтальное положение. Селезенка имеет форму уплощенной и удлиненной полусферы, темно-красного цвета (в связи с большим количеством эритроцитов, в ней депонирующихся), мягкой консистенции. Размеры селезенки у взрослого человека: длина – 10-14 см, ширина – 6-10 см, толщина – 3-4 см; масса – 150-190 г. У селезенки выделяют две поверхности:
диафрагмальную, обращена латерально и вверх (к диафрагме), гладкая и выпуклая
переднемедиальную (или висцеральную), обращена медиально и вниз (по направлению к органам брюшной полости), неровная, на ней имеются ворота селезенки (через посредство которых в селезенку входят селезеночные артерии и нервы, выходят селезеночные вены и лимфатические сосуды). Некоторые области этой поверхности селезенки имеют углубления, соответствующие местам ее контакта с дном желудка, левой почкой и левым надпочечником, левым изгибом ободочной кишки и некоторыми другими органами.
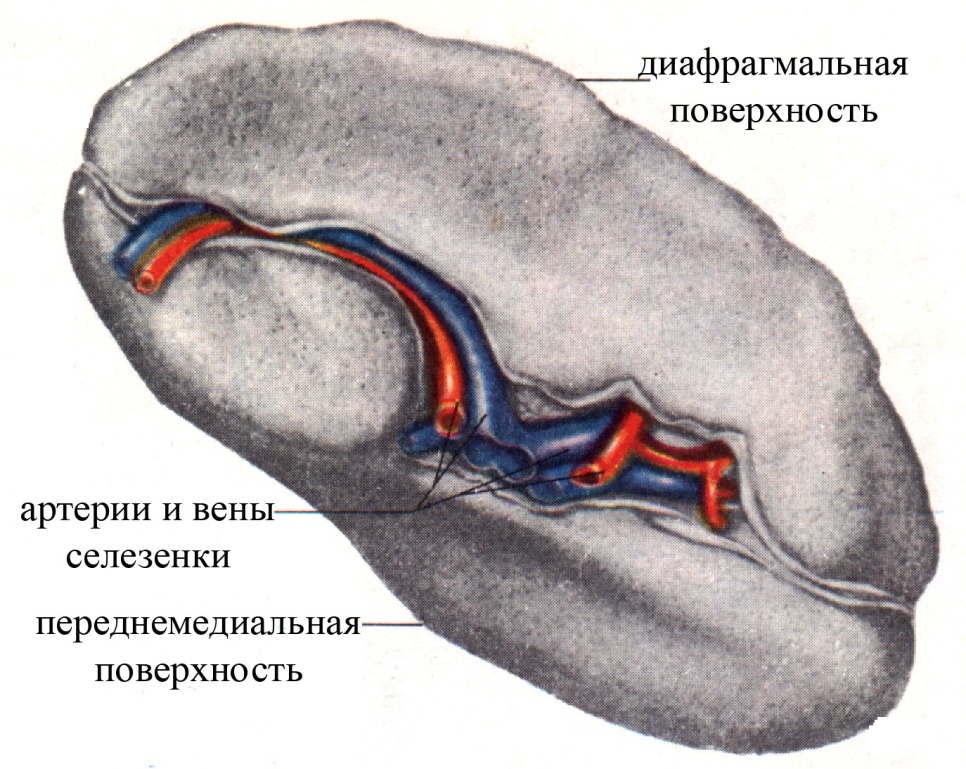
Рис. Селезенка (общий вид)
В эмбриональном периоде селезенка человека, наряду с красным костным мозгом, является органом гемопоэза. В постнатальном периоде она утрачивает эту функцию и представляет собой орган, в котором осуществляется:
антигензависимая пролиферация и дифференцировка лимфоцитов
очистка крови от антигенов
депонирование крови
элиминация дефектных и старых эритроцитов, и в меньшей степени – других клеточных элементов крови (кровяных пластинок и лейкоцитов)
обеспечивая элиминацию старых эритроцитов, селезенка тем самым принимает участие в обмене железа и образовании желчных пигментов. Так, железо, образующиеся в результате распада гемоглобина в селезеночных макрофагах, связывается с белком трансферрином и транспортируется в красный костный мозг, где используется для эритропоэза, а лишенный железа гемм гемоглобина превращается в билирубин (желчный пигмент) и транспортируется в печень, в составе желчи которой экскретируется в тонкий кишечник и в дальнейшем выводится из организма в составе кала
вырабатывает гуморальный фактор, тормозящий эритропоэз в красном костном мозге.
Несмотря на множественность функций селезенки, самой главной ее функцией, определяющей отношение к органам иммунной системы, является антигензависимая пролиферация и дифференцировка лимфоцитов (т.е. вторичный лимфоцитопоэз) и связанная с этим очистка крови, протекающей через селезенку, от различных антигенов. Причем селезенка, в отличие от лимфатических узлов, не получает лимфу по приносящим лимфатическим сосудам (лимфатические сосуды селезенки являются только выносящими), а, следовательно, не принимает участие в очистке лимфы, поступающей от других органов, но обеспечивает очистку притекающей крови от антигенов благодаря:
во-первых, длительному контакту крови с лимфоцитами селезенки вследствие депонирования крови в ней
во-вторых, наличию в ее паренхиме (называемой пульпой) большого количества лимфоцитов и антигенпредставляющих клеток (макрофагов и ретикулодендроцитов).
Г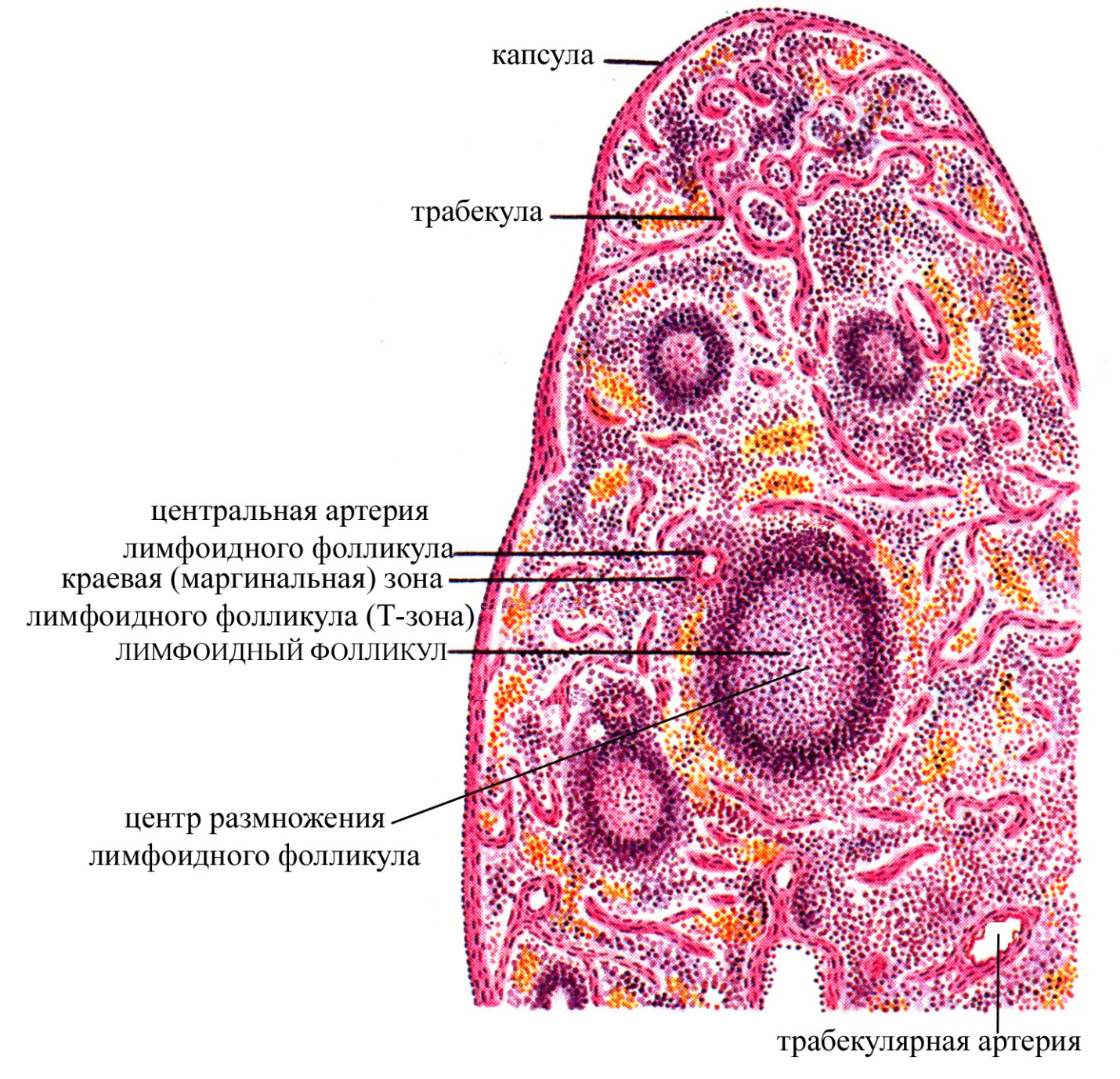 истоструктура
селезенки имеет некоторые особенности,
обусловленные ее функциональной
направленностью, в частности, необходимостью
обеспечивать как депонирование крови,
так и выброс ее в кровоток, а также
возможность фильтрации и очистки
протекающей крови от антигенов, старых
и дефектных эритроцитов и другие функции.
Так, подобно другим неполым органам,
селезенка покрыта соединительнотканной
капсулой, от которой внутрь органа
(преимущественно в области ворот)
отходят соединительнотканные трабекулы,
проходящие через паренхиму селезенки,
достигающие соединительнотканной
капсулы на противоположной воротам
выпуклой диафрагмальной поверхности
органа и формирующие в совокупности
соединительнотканную строму. Но в
отличие от большинства других неполых
органов, типичной особенностью селезенки
является наличие в ее капсуле и строме
большого количества эластических
волокон и гладкомышечных клеток, что
позволяет этим структурам растягиваться
в случае необходимости (при депонировании
крови в органе) и сжиматься, способствуя
выбросу крови в кровоток.
истоструктура
селезенки имеет некоторые особенности,
обусловленные ее функциональной
направленностью, в частности, необходимостью
обеспечивать как депонирование крови,
так и выброс ее в кровоток, а также
возможность фильтрации и очистки
протекающей крови от антигенов, старых
и дефектных эритроцитов и другие функции.
Так, подобно другим неполым органам,
селезенка покрыта соединительнотканной
капсулой, от которой внутрь органа
(преимущественно в области ворот)
отходят соединительнотканные трабекулы,
проходящие через паренхиму селезенки,
достигающие соединительнотканной
капсулы на противоположной воротам
выпуклой диафрагмальной поверхности
органа и формирующие в совокупности
соединительнотканную строму. Но в
отличие от большинства других неполых
органов, типичной особенностью селезенки
является наличие в ее капсуле и строме
большого количества эластических
волокон и гладкомышечных клеток, что
позволяет этим структурам растягиваться
в случае необходимости (при депонировании
крови в органе) и сжиматься, способствуя
выбросу крови в кровоток.
Рис. Гистоструктура селезенки (общий вид)
Паренхима селезенки, называемая пульпой, заполняет пространства между капсулой и трабекулами. В зависимости от особенностей гистологического строения пульпы и ее функциональной направленности, выделяют две ее разновидности:
белая пульпа, составляет примерно 1/5 от общего объема пульпы, занимает диффузное положение среди красной пульпы, группируясь преимущественно вокруг мелких артерий селезенки, вышедших из соединительнотканных трабекул в паренхиму органа. Имеет строение, типичное для лимфоидных органов: в основе ее лежит ретикулярная ткань, в ячеях которой находятся лимфоциты на разных стадиях антигензависимой дифференцировки и макрофаги. Белая пульпа селезенки представлена двумя типами структур:
периартериальными лимфоидными влагалищами, сопровождающими по ходу пульпарные артерии, берущие начало от артерий соединительнотканных трабекул
лимфоидными фолликулами, аналогичными таковым в лимфатических узлах и рыхлой волокнистой соединительной ткани.
Основной функцией белой пульпы является участие в иммунных реакциях клеточного и гуморального типа
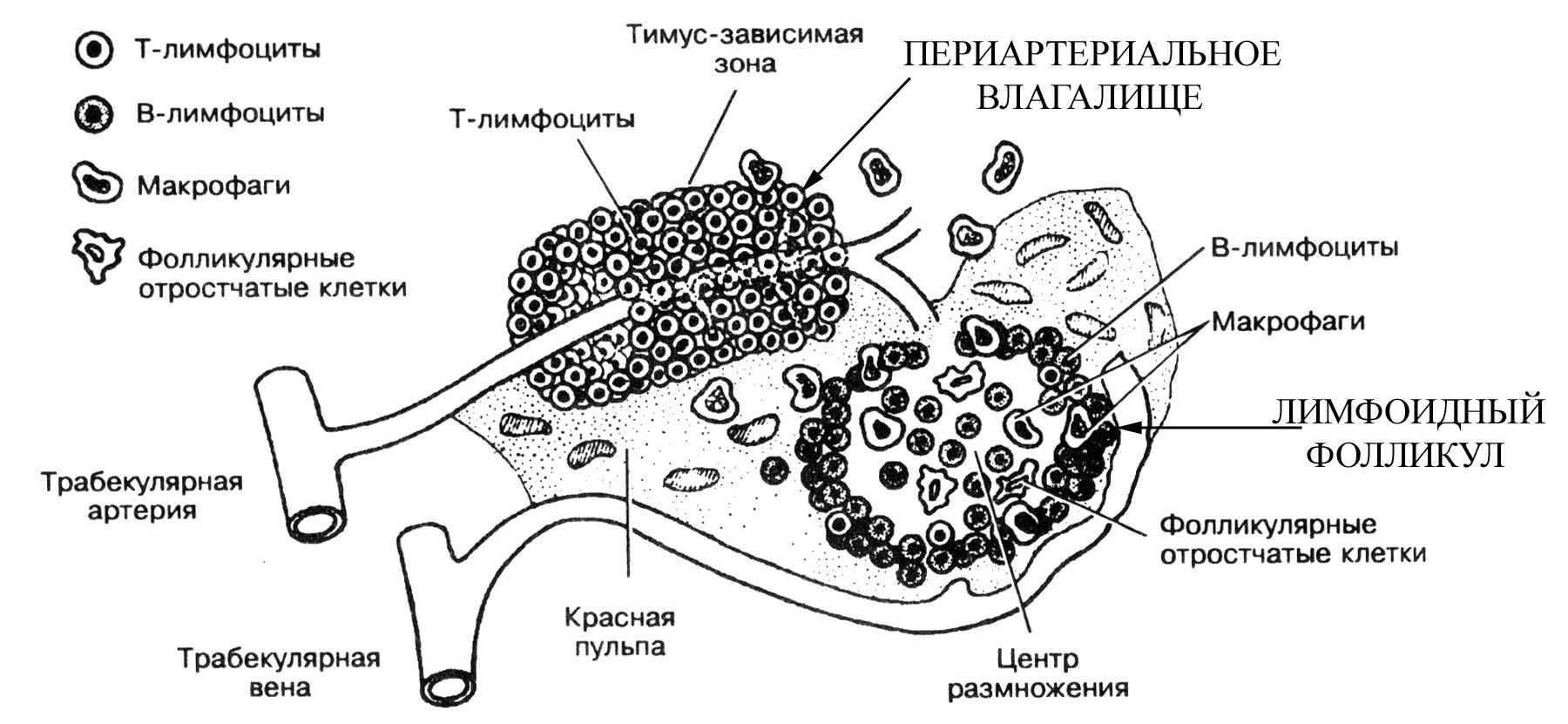
Рис. Схема локализации белой пульпы селезенки
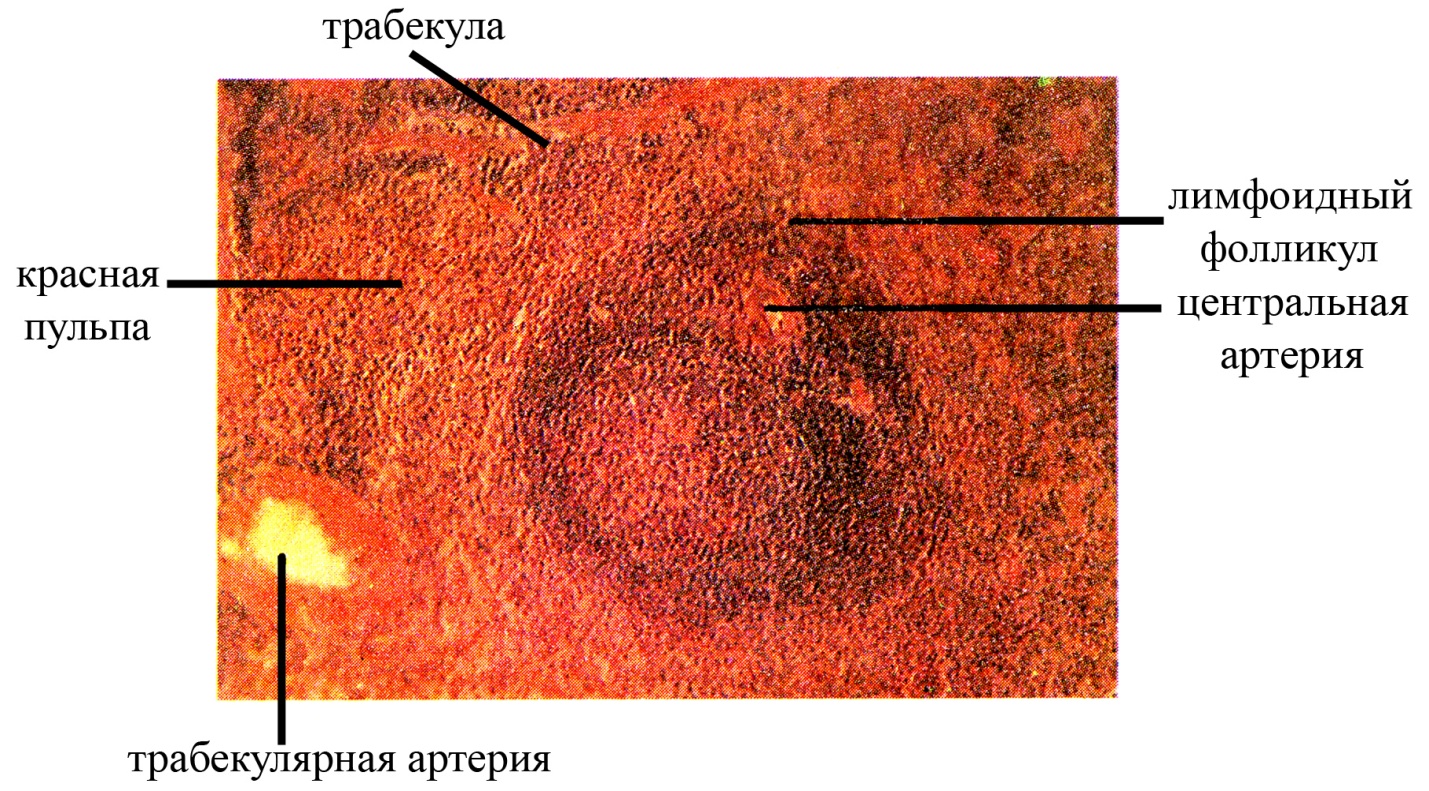
Рис. Лимфоидный фолликул селезенки (окраска гематоксилин-эозином)
красная пульпа, составляет 80-85% от общего объема пульпы селезенки, представлена т.н. пульпарными тяжами, между которыми проходят венозные синусоиды. В основе красной пульпы, подобно белой пульпе, лежит ретикулярная ткань, но в ячеях этой ткани содержаться преимущественно эритроциты, хотя присутствуют также и макрофаги, активированные лимфоциты и плазматические клетки, продуцирующие антитела. Ретикулиновые волокна красной пульпы связаны с коллагеновыми волокнами соединительнотканных трабекул и капсулы селезенки, образуя прочную сеть. Основной функцией красной пульпы является депонирование крови и элиминация из нее дефектных и старых клеточных элементов. Вместе с тем благодаря наличию в тяжах красной пульпы плазматических клеток, пришедших с током крови из белой пульпы, активированных Т- и В-лимфоцитов, приносимых с током крови из белой пульпы или из системного кровотока, она также принимает участие в иммунных реакциях (преимущественно гуморального типа).
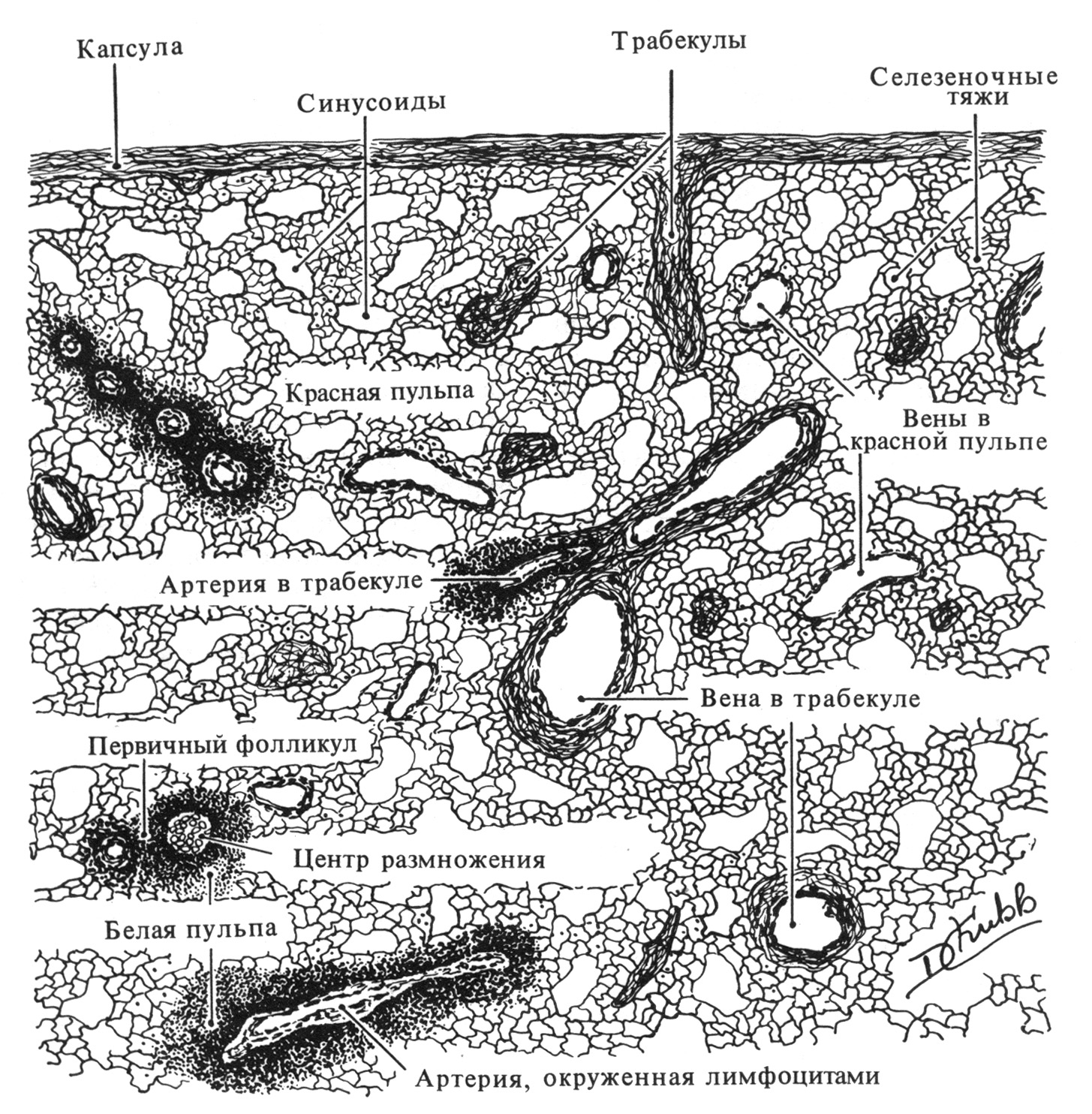
Рис. Схема, отражающая распределение красной и белой пульпы селезенки
Для полного понимания гистологического строения и функций селезенки, а также взаимосвязи между красной и белой пульпой необходимо ознакомиться с особенностями организации ее сосудистого русла. В области ворот к селезенке подходит селезеночная артерия, являющаяся ветвью аорты. Селезеночная артерия еще до вступления в ворота селезенки ветвится на несколько более мелких артерий, которые самостоятельно входят в орган. Эти артерии (называемые ветвями селезеночной артерии), войдя в соединительную ткань ворот селезенки, дают начало 4-5 сегментарным артериям. От сегментарных артерий ответвляются более мелкие артерии, которые проходят в составе соединительнотканных трабекул через всю паренхиму селезенки в направлении от переднемедиальной к диафрагмальной ее поверхности и называются трабекулярными. Трабекулярные артерии по своему ходу дают начало более мелким артериям, направляющимся от трабекул в пульпу селезенки и называемых в связи с этим пульпарными артериями (тонкие, их диаметр составляет в среднем 0,2 мм). Пульпарные артерии проходят в составе красной пульпы, поддерживаются сетью ретикулиновых волокон, в ячеях которой находятся лимфоциты (преимущественно Т-лимфоциты). Такие лимфоидные структуры, сопровождающие пульпарные артерии, называются периваскулярными (периартериальными) лимфоидными влагалищами. Наряду с Т-лимфоцитами в таких влагалищах присутствуют и макрофаги, а также обнаруживается небольшое количество В-лимфоцитов и плазматических клеток. Но в связи с тем, что преобладающими здесь являются Т-лимфоциты, эту зону называют тимусзависмой зоной селезенки (подобно тимусзависимой или паракортикальной зоне лимфатических узлов). Т-лимфоциты периартериальных влагалищ селезенки поступают сюда преимущественно из крови самих артерий, которые эти влагалища окружают, тогда как В-лимфоциты имеют тенденцию концентрироваться в лимфоидных фолликулах селезенки. Причины такой закономерности распределения Т- и В-лимфоцитов в селезенке не ясны. Лимфоциты периартериальных лимфоидных влагалищ (как активированные в результате контакта с определенным антигеном, так и неактивированные) могут переходить в красную пульпу, а оттуда поступать в кровоток (т.е. возвращаться в циркуляцию).
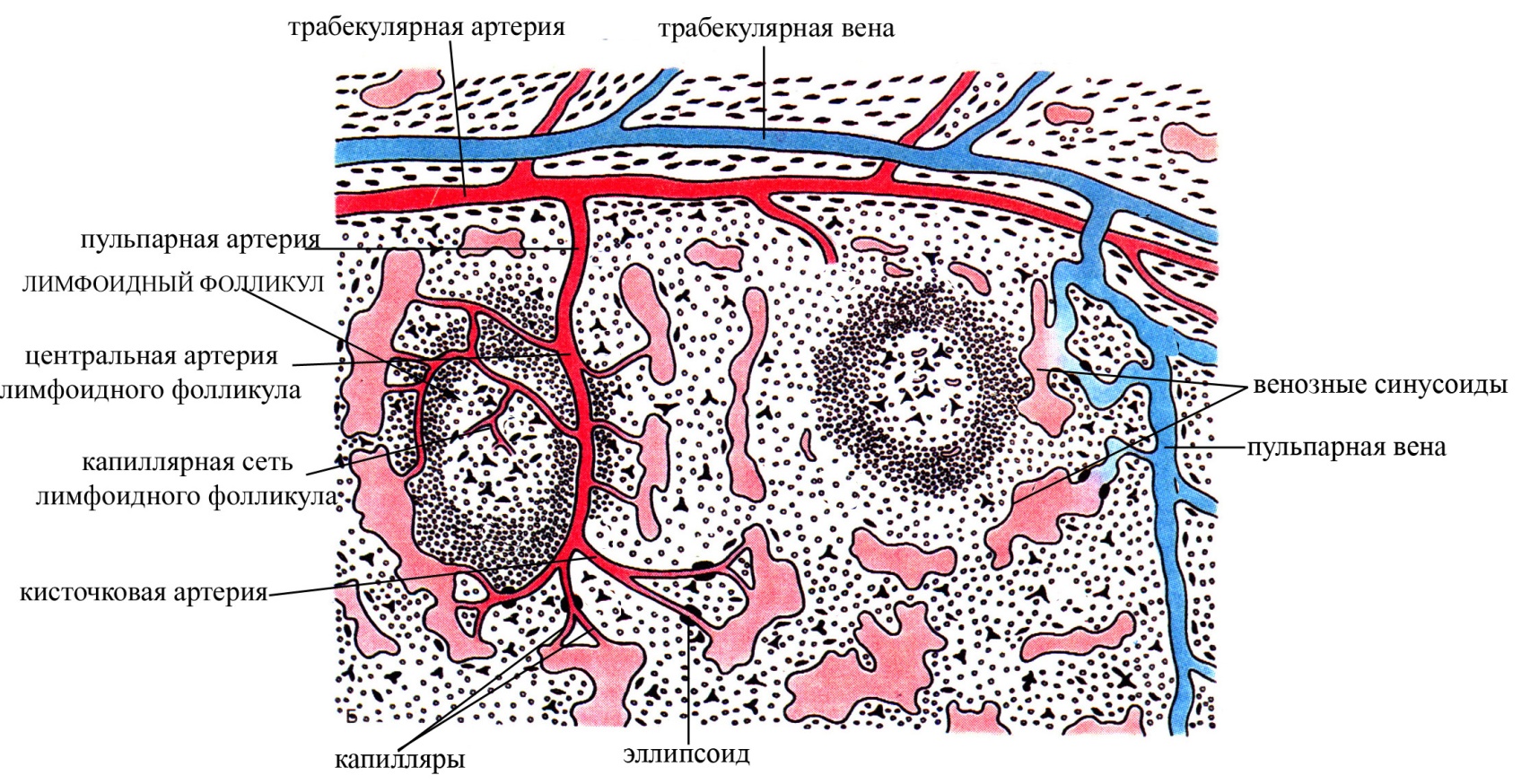
Рис. Схема кровоснабжения селезенки
В некоторых участках красной пульпы периваскулярные влагалища, сопровождающие пульпарные артерии, сильно расширяются и образуют типичные лимфоидные фолликулы, составляющие вместе с периартериальными лимфоидными влагалищами белую пульпу. Артерия, проходящая в лимфоидных фолликулах и являющаяся продолжением пульпарной артерии, называется центральной артерией и, по сути, является уже не истинной артерией, а артериолой, поскольку разветвляется на капиллярную сеть лимфоидного фолликула. Центральная артерия проходит в лимфоидном фолликуле, как правило, эксцентрично (в связи с тем, что она оттесняется к периферии фолликула центром размножения самого лимфоидного узелка). Центральная артерия лимфоидного фолликула, пройдя через него и дав начало его капиллярной сети, вновь выходит в красную пульпу, разветвляясь, подобно щетинкам кисточки на группу (включающую 2-6 ветвей) радиально расходящихся кисточковых артерий (по сути, тоже являются не истинными артериями, а артериолами, их диаметр составляет около 50 мкм). Кисточковые артерии, в свою очередь, разветвляются, как правило, на 2-3 мелкие артериолы, которые входят в т.н. эллипсоиды и образуют капиллярные сети самой красной пульпы. Эллипсоиды представляют собой сеть из ретикулярной ткани, содержащей скопления макрофагов и окружающей в виде муфт (гильз) мелкие артериолы. Такая муфта выступает в качестве своеобразного сфинктера на мелких артериолах, образующих капиллярные сети в красной пульпе селезенки.
Относительно того, куда открываются капилляры красной пульпы, до настоящего времени не существует однозначного мнения. Так, согласно представлениям одних исследователей (приверженцев теории замкнутой циркуляции в селезенке) капилляры красной пульпы открываются в венозные синусоиды, расположенные между ее тяжами. По мнению же других специалистов, придерживающихся теории незамкнутой циркуляции крови в селезенке, эти капиллярные сети красной пульпы открываются прямо в ее тяжи, откуда кровь поступает в венозные синусоиды, проходящие между ними. Наконец, существует и точка зрения, согласно которой допускается возможность излияния крови из капилляров как непосредственно в тяжи красной пульпы, так и в венозные синусоиды. Причем будет ли циркуляция в селезенке замкнутой или нет, зависит от степени растянутости органа. Так, в растянутой селезенке кровь из капилляров красной пульпы может переходить непосредственно в ее тяжи, а оттуда – в венозные синусоиды, тогда как в относительно спавшемся органе – капилляры открываются преимущественно в венозные синусоиды. Кровь из венозных синусоидов поступает в селезеночные вены (сначала пульпарные, затем трабекулярные), которые, объединяясь в более крупные вены, выходят из селезенки в области ворот и впадают в воротную вену печени.
Работами иностранных исследователей показано, что некоторые капилляры эллипсоидов красной пульпы открываются не в ее тяжи и не в синусоиды, а непосредственно в венулы, обеспечивая тем самым быстрый обходной путь крови в селезенке.
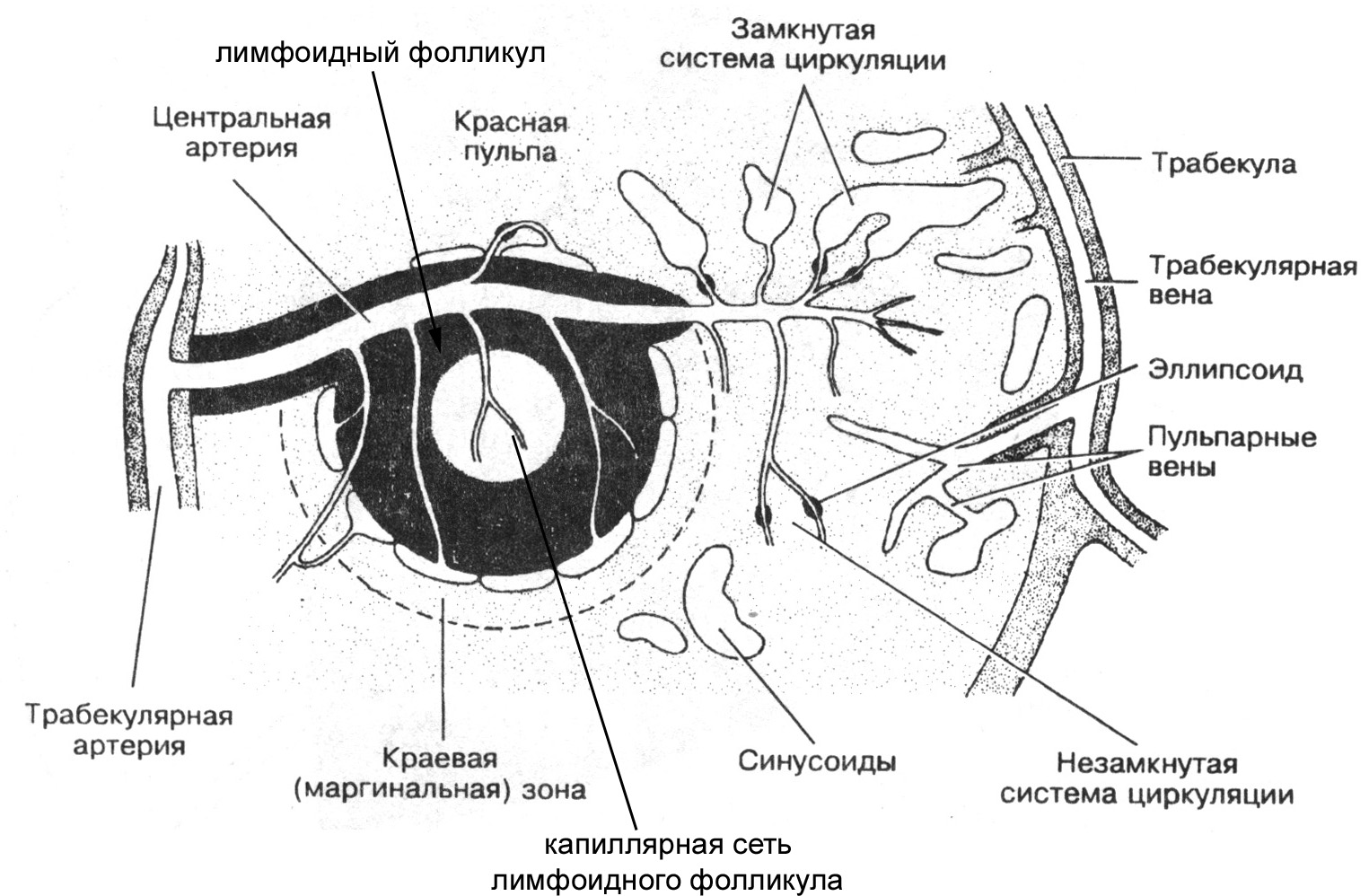
Рис. Схема, отражающая замкнутую и незамкнутую циркуляцию крови в селезенке
В енозные
синусоиды
красной пульпы селезенки представляют
собой каналы (диаметр просвета 12-40 мкм),
пронизывающие красную пульпу и окруженные
ее тяжами. Стенка этих синусоидов
образована удлиненными растянутыми
эндотелиальными клетками, расположенными
продольно; причем в растянутой селезенке
между этими клетками возникают широкие
продольные щели (синусоиды растянутой
селезенки похожи на старые бочонки,
продольные доски которых разошлись,
так что между ними возникли щели). Подобно
бочонкам, окруженным железными обручами,
эндотелиальные клетки венозных синусоидов
отчасти окружены и поддерживаются
"обручами" из ретикулиновых волокон,
связанных с материалом базальной
мембраны.
енозные
синусоиды
красной пульпы селезенки представляют
собой каналы (диаметр просвета 12-40 мкм),
пронизывающие красную пульпу и окруженные
ее тяжами. Стенка этих синусоидов
образована удлиненными растянутыми
эндотелиальными клетками, расположенными
продольно; причем в растянутой селезенке
между этими клетками возникают широкие
продольные щели (синусоиды растянутой
селезенки похожи на старые бочонки,
продольные доски которых разошлись,
так что между ними возникли щели). Подобно
бочонкам, окруженным железными обручами,
эндотелиальные клетки венозных синусоидов
отчасти окружены и поддерживаются
"обручами" из ретикулиновых волокон,
связанных с материалом базальной
мембраны.
Рис. Электронная микрофотография стенки венозного синусоида селезенки
На каждом конце синусоида (на входе и выходе из него) имеются сфинктеры, в зависимости от степени сокращения и расслабления которых синусоид может находиться в разных структурно-функциональных состояниях (т.н. фазах). Так, когда оба сфинктера открыты, синусоид относительно узкий и легко пропускает кровь (фаза пропускания). В случае закрытого выходного сфинктера при открытом входном синусоид наполняется кровью и обеспечивает эффективную ее фильтрацию (фаза фильтрации-наполнения); при этом стенки синусоида задерживают эритроциты, но позволяют плазме выходить в селезеночные тяжи, что в конечном итоге приводит к наполнению синусоида эритроцитами. Когда синусоид наполняется эритроцитами, входной сфинктер его закрывается (синусоид переходит в фазу запасания). При обоих открытых сфинктерах после фазы запасания синусоид переходит в фазу опорожнения, при которой сгустившиеся в нем эритроциты переходят в кровоток. Опорожнению венозных синусоидов и селезенки в целом, наряду с расслаблением сфинктеров самих синусоидов, способствует и сокращение гладкомышечных клеток капсулы и трабекул.
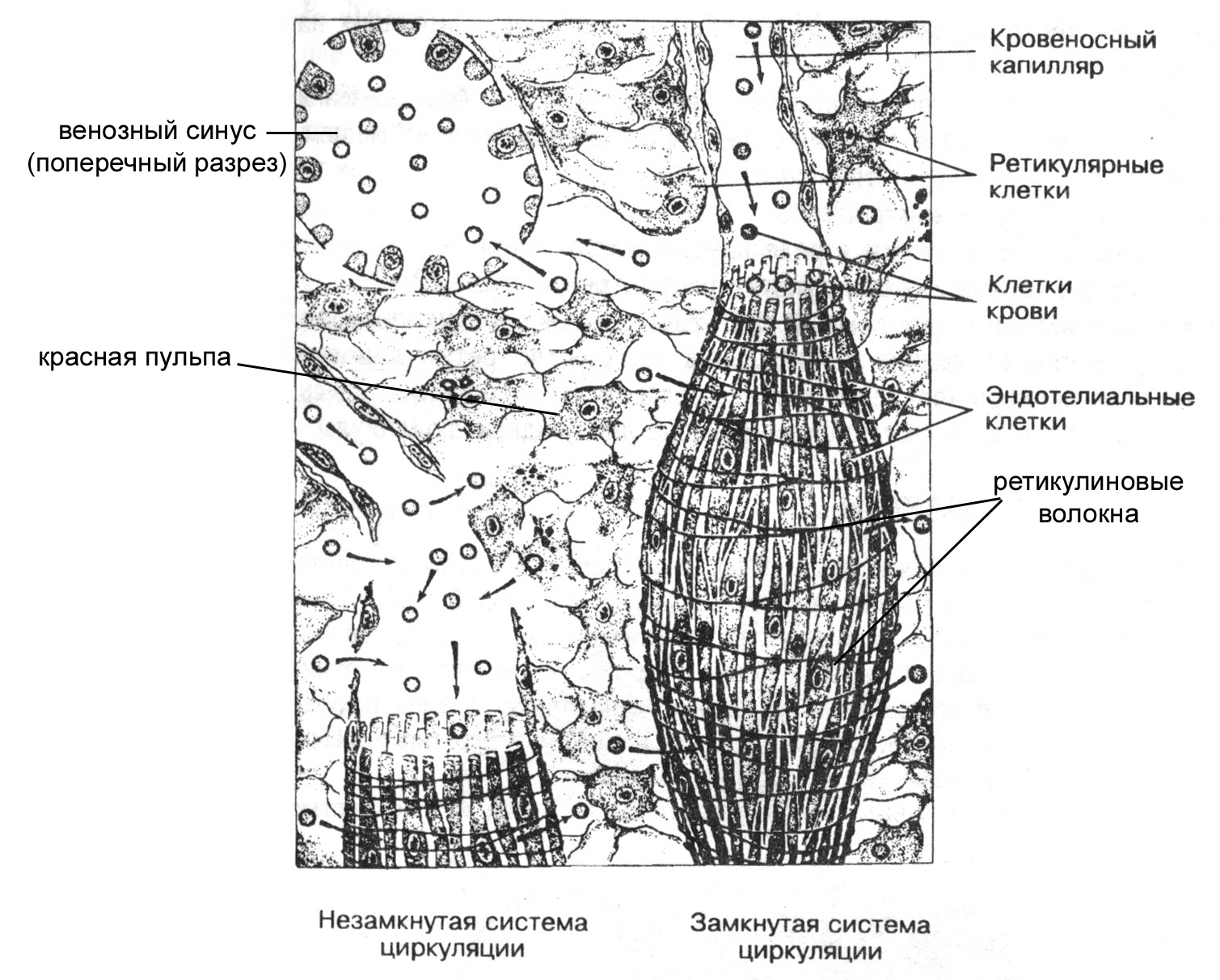
Рис. Схема строения венозного синуса селезенки
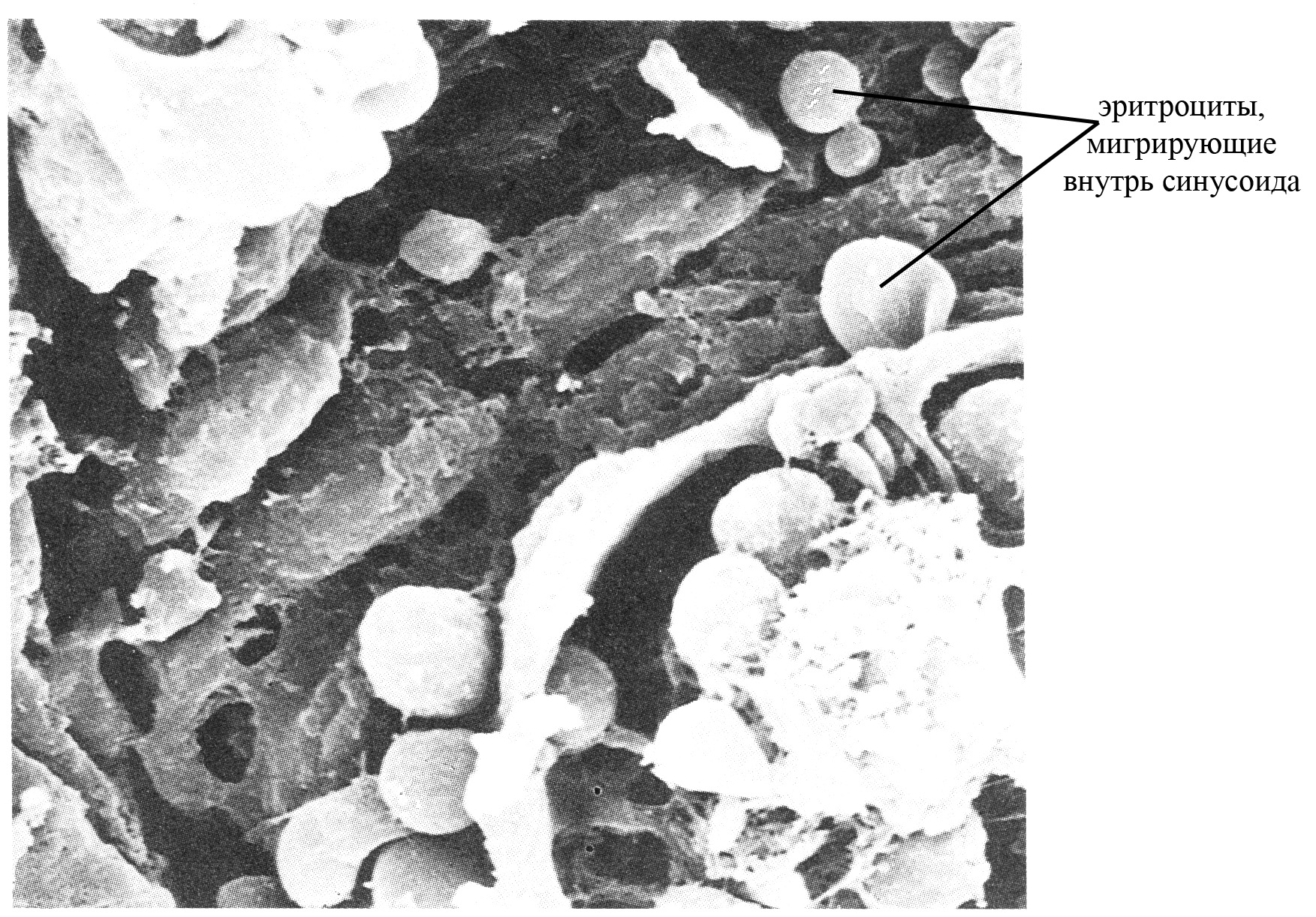
Рис. Миграция эритроцитов сквозь стенку венозного синусоида селезенки (сканирующая электронная микроскопия)
Эритроциты и отчасти лимфоциты из тяжей красной пульпы возвращаются в кровоток именно на уровне венозных синусоидов, чему способствуют широкие продольные щели в их стенках. Однако старые или какие-то дефектные эритроциты в связи с низкой своей эластичностью протиснуться через щели в стенках синусоидов не могут, деформируются и фагоцитируются макрофагами, присутствующими как в самой полости синусоидов, так и окружающими их стенку. Макрофаги красной пульпы, фагоцитировашие большое количество эритроцитов, частично поступают в селезеночную вену, по которой достигают печени и могут переходить в общий кровоток. С током крови они в конечном итоге поступают к легким и там, в связи со своими большими размерами, протиснуться через легочные капилляры не могут, переходят в легочные альвеолы, а затем отхаркиваются.
Селезенка начинает закладываться из мезенхимного зачатка на 5-6-й неделе внутриутробного развития и представляет собой плотное скопление мезенхимных клеток. Вскоре в мезенхимном зачатке образуются щели – будущие сосуды селезенки, вокруг которых осуществляется тканевая дифференцировка органа, из части мезенхимных клеток дифференцируется ретикулярная ткань, которая заселяется стволовыми клетками. На 2-4-м месяце внутриутробного развития в селезенке формируются венозные синусы и вокруг них появляются очаги гемопоэза (т.е. на данном этапе эмбрионального развития селезенка представляет собой типичный кроветворный орган). Максимальную интенсивность процессы миелопоэза в селезенке достигают на 5-м месяце внутриутробного развития. При этом уже в конце 4-го месяца эмбрионального развития в селезенке обнаруживаются скопления лимфоцитов, дающие начало в дальнейшем лимфоидным фолликулам и периартериальным влагалищам. К 8-му месяцу эмбриогенеза миелопоэз в селезенке уменьшается и в дальнейшем (к моменту рождения) прекращается вообще, а интенсивность лимфоцитопоэза, наоборот, нарастает.
В старческом возрасте белая и красная пульпа селезенки подвергается атрофии, количество лимфатических фолликулов и размеры центров размножения в них уменьшаются.