

Глава 4 патофизиология гипофиза Гормоны передней доли гипофиза (аденогипофиза)
К данной группе относятся гормоны семейства проопиомеланокортина, т.е. белка, который синтезируется в передней и средней долях гипофиза, а также в пептидергических нейронах гипоталамуса, и является "стволовой" молекулой (прогормоном), при расщеплении которой под действием специальных протеолитических ферментов (конвертаз) образуются: адренокортикотропный гормон (кортикотропин – АКТГ), α-, β- и γ-меланоцитстимулирующие гормоны (меланотропин – МСГ), β-липотропин, β-эндорфин. В передней доле гипофиза синтезируются также соматотропный гормон или гормон роста (соматотропин – СТГ, ГР), гонадотропные гормоны или гонадотропины (фолликулостимулирующий гормон – ФСГ; лютеинизирующий гормон – ЛГ), пролактин (лактогенный гормон), тиреотропный гормон (тиреотропин – ТТГ).
Секреция всех аденогипофизарных гормонов характеризуется суточной периодичностью. Как правило, в дневные часы уровень гормонов низкий, а в ночные часы возрастает.
Промежуточная доля гипофиза секретирует МСГ, задняя же доля депонирует и освобождает в кровоток нейрогормоны – вазопрессин и окситоцин, синтезируемые в нейросекреторных ядрах гипоталамуса – супраоптическом и паравентрикулярных.
Гормоны семейства проопиомеланокортина
Проопиомеланокортин (ПОМК) имеет 265 аминокислотных остатков. Тела ПОМК-содержащих клеток найдены не только в гипофизе, но и в нервной системе: в гипоталамическом аркуатном ядре, ядре солитарного тракта в каудальном отделе ствола головного мозга, в спинном мозге и спинномозговых ганглиях. Окончания их волокон определяются в различных областях гипоталамуса и в других мозговых структурах, в частности в лимбической системе.
В настоящее время известно, что многие ткани содержат клетки, способные продуцировать ПОМК, но на более низком уровне, чем в гипофизе, в том числе клетки кожи, селезенки и иммунокомпетентные клетки – лимфоциты и макрофаги.
Протеолитические ферменты, обеспечивающие внутриклеточное "разрезание" ПОМК (посттрансляционный процессинг) имеют тканевую и временную специфичность и обеспечивают образование гормонально активных пептидов. Принятая в настоящее время схема процессинга приведена на рис. 4.
Группа пептидов, имеющих сходные аминокислотные последовательности: АКТГ, α-, β- и γ-МСГ, – получили название меланокортиновых пептидов, или меланокортинов. Все они содержат неизменную последовательность четырех аминокислот: His-Phe-Arg-Trp, соответствующую фрагменту АКТГ4-9.
Биологические эффекты пептидов, происходящих из ПОМК, в большой степени опосредуются через меланокортиновые рецепторы (МК), пять типов которых описаны в настоящее время.
ПОМК
γ-МСГ
АКТГ
β-липотропин
β-эндорфин
β-МСГ
гипоталамус –
аденогипофиз –
соматостатин –
ГР
соматомедин
(инсулиноподобный
фактор роста –
ИПФР-1)
печень
+
ГРРГ
γ-липотропин
CLIP
α-МСГ




![]()
Рис. 4. Меланокортиновые пептиды: АКТГ и α-, β- и γ-МСГ, образующиеся при посттрансляционном процессинге ПОМК, являющегося также прекурсором для опиоидных пептидов и CLIP-кортикоподобного пептида промежуточной доли (по A. Catania, S. Gatti, G. Colombo, I. Lipton, 2004).
Хотя адреналовые стимулирующие эффекты АКТГ и пигментные влияния МСГ известны более 50 лет, исследования о том, что меланокортиновые пептиды обладают множественными эффектами в организме, являются более недавними. Эти эффекты несоизмеримы и простираются от модуляции лихорадки и воспаления до контроля приема пищи, автономных функций и экзокринной секреции. Более того, недавнее исследование показало, что определенные меланокортиновые пептиды обладают антимикробными эффектами.
В последние годы установлено, что посттрансляционное расщепление ПОМК в гипоталамусе может регулироваться в соответствии с энергетической потребностью. Эта регуляция опосредуется особым гормоном – лептином, представляющим собой циркулирующий в крови протеин. Он синтезируется в жировой ткани, что, в частности, подтверждается положительной корреляцией между концентрацией в ней мРНК лептина и процентом жира в теле.
Лептин обнаружен в цереброспинальной жидкости и может транспортироваться в мозг. Рецепторы к нему широко представлены в аркуатном ядре гипоталамуса.
В целом, лептин является частью петли обратной связи: жировая ткань продуцирует его как гуморальный сигнал, который пропорционален количеству жира и действует на гипоталамус, чтобы уменьшить потребление пищи и увеличить расход энергии. Сигналы лептина улавливаются и трансдуцируются ПОМК-ергическими нейронами: лептин прямо усиливает в них потенциалы действия. При этом происходящие из ПОМК биоактивные пептиды (АКТГ, МСГ) являются эндогенными лигандами МК-4 рецепторов, через которые осуществляется контроль аппетита и энергетического гомеостаза. Эта гипотеза получила название липостатической. Ее достоверность подтверждается целым рядом фактов. Так, у мышей с генетически отсутствующими рецепторами к лептину или МК-4 рецепторами проявляется фенотип с ожирением вследствие тяжелой гиперфагии. Уровни мРНК ПОМК значительно ниже в гипоталамусе ожиревших крыс с дефицитом рецепторов к лептину.
Эффекты АКТГ
Адренокортикотропное действие, т.е. стимуляция секреции коры надпочечников. Основной эффект – активация пучковой зоны и увеличение продукции кортизола, в меньшей степени – сетчатой зоны и секреции андрогенов и в малой степени – альдостерона.
Метаболическое действие. В основном зависит от продукции глюкокортикоидов, но включает и вненадпочечниковые эффекты.
Изменение белкового обмена при гиперпродукции АКТГ: АКТГ → усиление синтеза кортизола в надпочечниках → усиление катаболизма белков → в условиях повышенного распада белков – усиление анаболизма.
Влияние избытка АКТГ на жировой обмен:
а) АКТГ → вненадпочечниковый механизм: усиление липолитической активности жировой ткани → усиление распада жиров → выход в кровь свободных жирных кислот и их окисление до кетоновых тел;
б) АКТГ → надпочечниковый механизм: торможение мобилизации жира + торможение липолитического действия СТГ → активация глюконеогенеза → стимуляция образования жира.
Нейротропное действие обусловлено N-концевым фрагментом молекулы гормона – АКТГ4-10 (4-10 – обозначение последовательности аминокислотных остатков). Основные эффекты: улучшение внимания и кратковременной памяти, ускорение обучения человека и животных, улучшение выбора биологически целесообразного поведения при стрессе.
Влияние на образование меланина и пигментацию кожи. Действие аналогично эффекту МСГ и объясняется тем, что АКТГ содержит в своей молекуле всю аминокислотную последовательность меланоцитстимулирующего гормона.
Патология
Гиперсекреция АКТГ наблюдается при базофильной аденоме передней доли гипофиза и нарушениях в гипоталамусе (недостаток дофамина) и приводит к развитию болезни Иценко-Кушинга. Основной механизм патогенеза – избыточная стимуляция под действием АКТГ коры надпочечников и развитие вторичного гиперкортизолизма. Гиперпродукция кортизола проявляется характерными нарушениями метаболизма, связанными с повышенным распадом белков, глюконеогенезом, перераспределением липидов. Однако интенсивность этих проявлений меньше, чем при первичном гиперкортизолизме, вследствие анаболических эффектов андрогенов, секреция которых также стимулируется избытком АКТГ. У женщин при этом в значительной степени проявляются вирилизм, гирсутизм, нарушение менструаций. Схемы соотношений в гипоталамо-гипофизарно-адреналовой оси в норме и при болезни Иценко-Кушинга представлены на рис. 5 и 6.
Лечение болезни Иценко-Кушинга путем двусторонней адреналэктомии приводило к развитию синдрома Нельсона, который проявляется гиперпигментацией кожных покровов и слизистых оболочек и хронической недостаточностью надпочечников. Нарушение обратной связи приводит к разрастанию аденомы и дальнейшей гиперсекреции АКТГ, с чем связывают усиление его меланоцитстимулирующего эффекта.
Меланотропины
α- и β-МСГ действуют на пигментные клетки (меланоциты). Вызывают распределение пигмента вдоль островков клеток, вследствие чего кожа темнеет. При недостатке гормона глыбки пигмента концентрируются в центре клетки – кожа светлеет. Гормон участвует в темновой адаптации глаза, повышает остроту зрения.
Установлено также нейротропное действие МСГ: улучшение внимания, восприятия и кратковременной памяти.
Кроме того, следует отметить антипиретические эффекты меланокортинов. Так, α-МСГ ослаблял лихорадку при центральном введении в дозах, которые не оказывали влияния на нормальную температуру тела. АКТГ1-24 и АКТГ1-39, содержащие такую же последовательность аминокислот, также ослабляли лихорадку. Эффект сохранялся у адреналэктомированных животных.
Эти эффекты α-МСГ и его присутствие в мозге позволили предположить, что этот пептид может быть эндогенным модулятором лихорадки. В экспериментах на животных установлено, что во время лихорадки увеличивается его содержание в медиобазальном гипоталамусе и перегородке. Инактивация же пептида, образующегося в естественных условиях, ослабляла лихорадку, но не влияла на нормальную температуру тела.
Антипиретические эффекты α-МСГ опосредуются меланокортиновыми рецепторами 4-го типа.
ЦНС
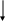

гипоталамус
аденогипофиз
АКТГ
кортизол
КРГ














Р ис. 5. Схема соотношений в гипоталамо-гипофизарно-адреналовой оси в норме. Условные обозначения:
– активирующие влияния,
– угнетающие влияния.
Липотропины
Липотропин (β- и γ-) обладает липотропной активностью, а также некоторой кортикотропной и меланоцитстимулирующей активностью. Механизм липотропного действия: активация липазы жировой ткани → липолиз и освобождение жирных кислот. Кроме того, липотропины ускоряют обмен глюкозы в жировой ткани.
Эндорфины
Эндорфины относятся к группе опиоидных пептидов, рецепторы которых обнаружены, в первую очередь, в мозговых структурах, имеющих непосредственное отношение к проведению болевых импульсов и восприятию боли (околоводопроводное серое вещество среднего мозга, лимбическая система, спинной мозг). Эндорфины и другие опиоидные пептиды – энкефалины - обнаружены в различных отделах мозга.
ЦНС

гипоталамус
АКТГ ↑
кортизол ↑
КРГ
аденогипофиз













Рис. 6. Изменения соотношения между компонентами гипоталамо-гипофизарно-адреналовой оси при болезни Иценко-Кушинга, вызванной аденомой аденогипофиза.
Утолщенные линии обозначают усиление влияний и функции, пунктирные – ослабление влияний. Остальные обозначения как на рис. 5.
Биологические эффекты:
Выполнение функции медиаторов анальгетической (антиноцицептивной системы.
Влияние на эмоциональное состояние – формирование чувства удовлетворения, успокоения (поэтому эти пептиды называют пептидами вознаграждения), а также радостного, приподнятого настроения – эйфории.
Влияние на обучение и память.
Модулирующее действие на развитие иммунного ответа.
Участие в регуляции секреции пролактина, СТГ, ЛГ.
Стресс-лимитирующее действие.
Соматотропный гормон
Соматотропный гормон (СТГ), или гормон роста (ГР) – пептидный гормон, содержащий 191 аминокислотный остаток.
Особенности механизмов действия на ткани. На основании сопоставления метаболического действия СТГ in vivio и in vitro было сделано предположение об опосредованном участии его эффектов специальными медиаторами, образующимися в организме. В дальнейшем было установлено существование таких посредников, названных соматомединами. В настоящее время известно 4 соматомедина: А, А1, В и С. Они представляют собой полипептиды, синтезируемые в печени. Однако не исключается возможность их продукции в других тканях.
Соматомедины (в первую очередь соматомедин С) вовлечены в регуляцию секреции СТГ по механизму отрицательной обратной связи. В торможении секреции гормона принимает участие соматостатин, продуцируемый в гипоталамусе.
Эффекты СТГ
В настоящее время эффекты СТГ разделяют на:
1) прямые: стимуляция синтеза соматомединов, в первую очередь, - соматомедина С (синоним – инсулиноподобный фактор роста-1 или ИФР-1) в печени и других тканях, стимуляция липолиза в жировой ткани и продукции глюкозы в печени;
2) непрямые: анаболическое действие и стимуляция роста. Эти эффекты опосредуются ИФР-1 и подавляются глюкокортикоидами.
Схема регуляции выделения ГР представлена на рис. 7.
Из рисунка видно, что секреция ГР стимулируется ГР-рилизинг гормоном (ГРРГ) и угнетается соматостатином. Отрицательная обратная связь на уровне гипофиза осуществляется соматомедином С (инсулиноподобный фактор роста-1). Этот фактор действует также на уровне гипоталамуса и стимулирует секрецию соматостатина.
Кроме этого, на основе фармакологических данных установлено, что выделение ГРРГ стимулируется ацетилхолином, α-адренергическими и допаминергическими влияниями, но угнетается β-адренергическими.
Секреция соматостатина (in vitro) стимулируется ацетилхолином и VIP и угнетается ГАМК.
В естественных условиях выделение ГР модифицируется эндогенными ритмами сна, стрессом и физическими упражнениями.
Механизмы эффектов