

Вопрос 3(25)Структура популяции:возростная,половая,экологическая.
http://ggf.bsu.edu.ru/ElBook/Ekologia/start.htm посмотреть тут!!!
СТРУКТУРА ПОПУЛЯЦИИ
характер распределения особей в пространстве, а также по половым, возрастным и другим морфологическими и физиологическим признакам. Пространственная структура популяции характеризует обычно типы распределения особей в биотопах (дисперсию) и в свою очередь делится на равномерное, агрегированное и случайное распределение особей. По иерархической структуре популяции (Наумов, 1963) различают географические, локальные (местные или экологические), элементарные (микропопуляции) и парцеллярные (парцеллы) популяции. Демографическая структура популяции сводится к возрастному и половому составу. Большинство популяций животных характеризуется также определенной социальной структурой популяции. Рассмотрим более подробно такие структуры популяции ,как возрастная ,половая,экологическая. (http://dic.academic.ru/dic.nsf/ecolog/7037/%D0%A1%D0%A2%D0%A0%D0%A3%D0%9A%D0%A2%D0%A3%D0%A0%D0%90)
4.3. Половая структура популяций Половая структура популяции определяет соотношение в ней особей разного пола. Генетический механизм определения пола обеспечивает расщепление потомства по полу в отношении, близком 1:1. Это так называемое первичное соотношение полов. Оно определяется генетическими механизмами - равномерностью расхождения половых хромосом. Например, у человека ХУ-хромосомы определяют развитие мужского пола, а XX - женского. В этом случае первичное соотношение полов 1:1, т.е. равновероятно. Однако в популяции соотношение полов, как правило, не равно 1 : 1 (количество самцов и самок различно). Это связано с тем, что сцепленные с полом признаки часто определяют значительные различия в физиологии, экологии и поведении самцов и самок. В связи с этим в популяциях часто наблюдается более высокая вероятность гибели представителей какого-либо пола, и значит, соотношение полов в популяции изменяется. Такое соотношение полов при рождении и на ранних стадиях развития организмов, когда на генетическую обусловленность накладываются различные экологические факторы, называется вторичным соотношением. Оно может существенно отличаться от первичного по целому ряду причин: избирательностью яйцеклеток к сперматозоидам, несущим X- или У-хромосому, неодинаковой способностью таких сперматозоидов к оплодотворению, различными внешними факторами. Например, зоологами описано влияние температуры на вторичное соотношение полов у рептилий. Аналогичная закономерность характерна и для некоторых насекомых. Так, у муравьев оплодотворение обеспечивается при температуре выше 20 °С, а при более низких температурах откладываются неоплодотворенные яйца. Из последних вылупляются самцы, а из оплодотворенных - преимущественно, самки. У некоторых видов пол изначально определяется не генетическими, а экологическими факторами. Например, корнеплод из семейства ароидных аризема японская дает растения с женскими цветами только из самых крупных и хорошо развитых клубней. Из мелких и слабых клубней образуются растения с мужскими цветами. У некоторых животных (например, амфибий) такие факторы среды, как температура, содержание и концентрация гормонов могут влиять на развитие таким образом, что половые различия будут отличаться от первичного хромосомного набора. Так, в среднеевропейских популяциях травяной лягушки облик молодых животных отражает внешность самок. Лишь к концу второго года жизни половина из них превращается в самцов. У муравьев, пчел и других общественных насекомых число цариц (самок, способных к половому размножению) в популяции регулируется рабочими особями посредством специфического питания. Соотношение полов среди половозрелых, размножающихся особей, называется третичным соотношением. Половой состав на данной стадии развития организмов весьма динамичен и изменчив. Для человека и других млекопитающих отмечена общая закономерность, заключающаяся в уменьшении доли самцов в старших возрастных группах. Например, у человека вторичное соотношение полов составляет 100 девочек на 106 мальчиков. По достижении 18- летнего возраста это соотношение выравнивается из-за повышенной мужской смертности. К 50 годам третичное соотношение составляет уже 85 мужчин на 100 женщин, а к 80 годам - 50 мужчин на 100 женщин. Следует отметить, что половая структура, т.е. соотношение полов, имеет прямое отношение к воспроизводству популяции и ее устойчивости.( ОСНОВЫ ЭКОЛОГИИ (В.В. МАВРИЩЕВ) 3-е изд., испр. и доп. - Минск: Выш. шк., 2007. - 447 с.) |
Половая структура отражает определенное соотношение мужских и женских особей в популяции. Генетический механизм определения пола обеспечивает расщепление потомства по полу в соотношении 1: 1. В силу разной жизнеспособности мужских и женских особей это первичное соотношение полов при оплодотворении часто заметно отличается от вторичного (при рождении — у млекопитающих) и тем более от третичного, характерного для половозрелых особей. Например, в популяциях человека вторичное соотношение полов составляет 100 девочек/106 мальчиков; к 16—18 годам это соотношение выравнивается и становится равным 1:1, к 50 годам— 100 женщин/85 мужчин, а к 80 годам соотношение по полу становится 2:1 (100 женщин/ 50 мужчин).
Изменение половой структуры популяции отражается на ее роли в экосистеме, так как самцы и самки многих видов отличаются друг от друга по характеру питания, ритму жизни, поведению и др. Так, самки некоторых видов комаров, клещей и мошек являются кровососущими, в то время как самцы питаются соком растений или нектаром. Преобладание доли самок над самцами обеспечивает более интенсивный рост популяции.( http://sbio.info/page.php?id=161)
ВОЗРОСТНАЯ СТРУКТУРА ПОПУЛЯЦИЙ
Важной характеристикой при исследовании популяции является ее возрастная структура, влияющая как на рождаемость, так и на смертность. Соотношение разных возрастных групп в популяции определяет ее способность к размножению и показывает ее перспективу. В быстрорастущих популяциях молодые особи составляют большую долю. Поэтому состояние популяции по прошествии определенного промежутка времени будет зависеть от ее нынешнего полового и возрастного состава. Возрастной состав популяции наглядно отражают так называемые возрастные пирамиды, представляющие собой гистограммы возрастного распределения особей. Если в популяции размножение происходит постоянно, то по возрастной структуре устанавливают - сокращается или увеличивается численность. В большинстве популяций репродуктивная способность их членов изменяется с возрастом. В современной экологии при исследовании возрастного состава популяции выделяют три экологические возрастные группы: - пререпродуктивную; - репродуктивную; - пострепродуктивную. Длительность этих возрастов по отношению к общей продолжительности жизни сильно варьирует у разных организмов. Как правило, наибольшей жизнеспособностью отличаются популяции, в которых все возрасты представлены относительно равномерно. Такие популяции называют нормальными. Если в популяции преобладают старые особи, это однозначно свидетельствует о наличии отрицательных факторов в ее существовании, нарушающих воспроизводительные функции. Такие популяции рассматривают как регрессивные, или вымирающие. Требуются срочные меры по выявлению причин такого состояния и их исключению. Популяции, представленные в основном молодыми особями, рассматриваются как внедряющиеся, или инвазионные. Жизненность их обычно не вызывает опасений, но велика вероятность вспышек чрезмерно высокой численности особей, поскольку в таких популяциях не сформировались трофические и другие связи. Особенно опасно, если такие популяции представлены видами, которые здесь ранее отсутствовали. В таком случае эти популяции обычно находят и занимают свободную экологическую нишу и реализуют свой потенциал размножения, интенсивно увеличивая численность. При благоприятных условиях в популяции имеются все возрастные группы и поддерживается сравнительно стабильный уровень численности. На возрастной состав популяции, помимо общей продолжительности жизни, влияют длительность периода размножения, число генераций в сезон, плодовитость и смертность разных возрастных групп. Например, у полевок взрослые особи могут давать потомство 3 раза в год или более, а молодые особи способны размножаться через 2-3 месяца. В популяциях растений иногда наблюдается еще более сложная картина. Дубы, например, дают семенную продукцию в течение нескольких столетий. Как результат, популяции у них формируются из огромного количества возрастных групп. Если популяция находится в нормальном или близком к нормальному состоянии, человек из нее может изымать то количество особей или биомассу (последний показатель обычно используется применительно к растительным сообществам), которая прирастает за промежуток времени между изъятиями. Ясно, что изыматься должны прежде всего особи послепродуктивного возраста (окончившие размножение). Если преследуется цель получения определенного продукта, то возраст, пол или другие характеристики популяций корректируются с учетом поставленной задачи. Обычно в начальный период роста (пререпродуктивная стадия) организмы размножаться не способны. Длительность этого периода у различных видов сильно варьирует - от нескольких минут у микроорганизмов до нескольких лет у человека, многих млекопитающих, деревьев. Пререпродуктивный период может продолжаться большую часть жизни, как, например у поденок (личиночное развитие в воде занимает от года до нескольких лет из-за длительного развития личинок) и 17-летней цикады (пререпродуктивная стадия достигает нескольких лет). Однако характерно, что репродуктивный период у этих видов очень короток (у поденок несколько дней, у цикады менее одного сезона), а пострепродуктивный период и вовсе практически отсутствует, как у многих других видов.
Рис. 4.1. Соотношение продолжительности пререпродуктивного (1), репродуктивного (2) и пострепродуктивного (3) периодов онтогенеза у некоторых видов Иллюстрацией вышесказанному является рис. 4.1, на котором показано соотношение пререпродуктивного, репродуктивного и пострепродуктивного периодов у разных видов беспозвоночных и позвоночных животных. Видно, что у поденок и сосновой совки пререпродуктивный период составляет более 95 % общей продолжительности жизни. У саранчи же и дрозофилы пререпродуктивный период не превышает 50 % общей продолжительности жизни. Иное положение наблюдается в популяциях человека, а также животных, которые содержатся в искусственно созданных условиях (комнатные, домашние животные, обитатели зоопарков). Особи в таких популяциях доживают до пострепродуктивного периода. У современного человека три эти «возраста» примерно одинаковы, на каждый из них приходится около трети жизни. У первобытных людей пострепродуктивный период был намного короче. В настоящее время соотношение возрастных экологических групп в популяции людей меняется. Увеличивается число детей, подростков и пенсионеров, т.е. непроизводительных слоев населения. Доля детей до 15 лет в большинстве развивающихся стран увеличилась до 50 %, пожилых людей старше 65 лет - до 15 %. Такое изменение соотношения возрастных групп приводит к увеличению нагрузки на трудоспособную часть населения. .( ОСНОВЫ ЭКОЛОГИИ (В.В. МАВРИЩЕВ) 3-е изд., испр. и доп. - Минск: Выш. шк., 2007. - 447 с.) |
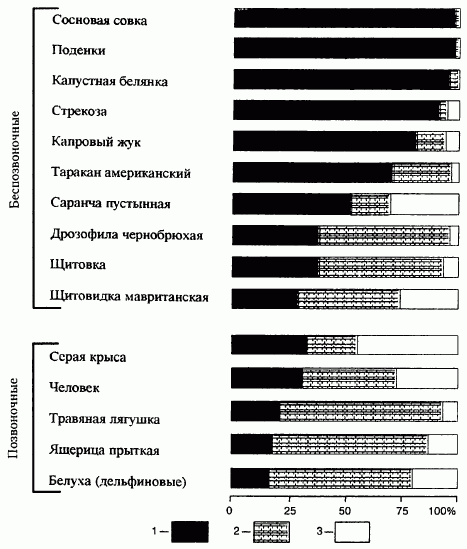
Возрастная структура отражает соотношение различных возрастных групп в популяциях, зависящее от продолжительности жизни, времени наступления половой зрелости, числа потомков в помете, количества потомств за сезон и др. Если какая-либо возрастная группа сокращается либо увеличивается, это сказывается на общей численности популяции. Например, массовое истребление крупных половозрелых особей в результате промысла приводит к резкому снижению численности популяции вследствие слабого пополнения ее молодыми особями. Поэтому присутствие в популяции большого количества особей младших возрастных групп свидетельствует о ее благополучии. Если же в популяции преобладают старые особи, можно со всей определенностью сказать, что данная популяция завершает свое существование.
Экологическая структура свидетельствует об отношении различных групп организмов к условиям окружающей среды. Например, особи одной популяции растений различаются рядом признаков: по размерам, количеству побегов, цветков, плодов, семян и т. п. Кроме того, разные особи этой же популяции зацветают неодновременно, что способствует более полному их опылению (при одновременном и кратковременном цветении насекомые могут не успеть опылить все цветки). У такой популяции меньший риск остаться без семян, например в случае кратковременных заморозков (замерзнет лишь часть цветков).
Динамика популяций. Теоретически любая популяция способна к неограниченному росту численности, если ее не лимитируют факторы внешней среды (ограниченность ресурсов, болезни, хищники и т. п.). В таком гипотетическом случае скорость роста популяции будет зависеть только от величины биотического потенциала, свойственного каждому конкретному виду. Биотический потенциал отражает теоретически возможное число потомков от одной пары (или одной особи) за определенный промежуток времени, например за весь жизненный цикл или за год.
У разных видов величина биотического потенциала резко различается. Например, у крупных млекопитающих даже при самых благоприятных условиях численность может возрастать лишь в 1,05—1,1 раза за год. У многих насекомых и ракообразных (тли, дафнии) численность за год увеличивается в 1010—1030 раз, а у бактерий еще больше. Однако в этих случаях в идеальных условиях численность любой популяции за определенное время будет расти в геометрической прогрессии (рис. 14.2). Рост численности с постоянной скоростью называется экспоненциальным ростом. Кривая, отражающая на графике подобный рост популяции, быстро увеличивает крутизну и уходит в бесконечность.
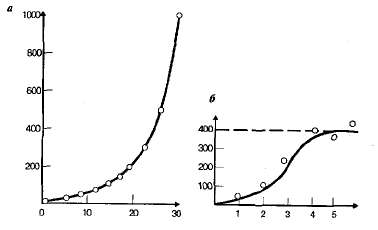
Рис. 14.2. Экспоненциальная (теоретическая) (а) и логистическая (реальная) (б) кривые роста популяций.
В природных условиях экспоненциальный рост популяций наблюдается крайне редко. Например, он был отмечен для популяции кролика, завезенного а Австралию, где для него имелись неограниченные пищевые и пространственные ресурсы при отсутствии сдерживающего влияния хищников. Такой тип роста наблюдается также при вспышках численности саранчи, непарного шелкопряда и других насекомых. Однако периоды экспоненциального роста обычно кратко временны.
При увеличении плотности популяции обычно наблюдается замедление роста численности, поскольку популяция оказывается в условиях с ограниченными ресурсами. Например, животным при высокой плотности популяции может не хватать пищи, а растения начинают затенять друг друга или им недостает влаги. Тип роста популяции при ограниченных ресурсах, характеризующийся снижением скорости по мере увеличения плотности популяции, называется логистическим (см. рис, 14.2).
Общие изменения численности популяции определяются такими процессами, как рождаемость, смертность и миграция особей.
Рождаемость характеризует частоту появления новых особей в популяции. Средняя величина рождаемости каждого вида определилась исторически как приспособление для восполнения убыли популяции. Различают абсолютную и удельную рождаемость.
Абсолютная рождаемость — количество особей, родившихся (вылупившихся, отпочковавшихся и т. д.) в популяции за единицу времени. Удельная рождаемость — количество особей, родившихся в популяции за единицу времени в расчете на одну особь. Показатель удельной рождаемости позволяет сравнивать скорость рождаемости в популяциях с разной численностью.
Величина рождаемости тем выше, чем больше доля особей, принимающих участие в размножении, чем выше плодовитость, чем чаще следуют друг за другом репродуктивные циклы. Обычно рождаемость в каждой популяции уравновешена характерной для нее смертностью.
Смертность характеризует скорость убывания численности популяции вследствие гибели особей от хищников, болезней, паразитов, генетической и физиологической неполноценности, старости и т. п.
На численность популяций существенное влияние оказывает также миграция особей.
Миграции — закономерные перемещения животных между существенно различными, пространственно разобщенными средами обитания. Подобные переселения вызываются изменением условий существования в местах обитания или изменением требований животного к этим условиям на разных стадиях развития. Массовое перемещение особей между популяциями может изменить их структуру и основные свойства (предотвратить гибель популяции, находящейся на грани вымирания, или, наоборот, привести ее к резкому сокращению). Например, массовое выселение оседлых животных (белок, кедровок, леммингов и др.) при внезапном ухудшении условий (засухи, пожары, наводнения и т. п.), перенаселении часто заканчивается их массовой гибелью.
Миграции (суточные, сезонные) позволяют организмам использовать оптимальные условия среды в таких местах, где их постоянное проживание невозможно. Они приводят к освоению новых биотопов, расширению общего ареала вида, к обмену особей между популяциями, увеличивают единство и общую устойчивость вида, способствуют успеху в борьбе за существование.
При отсутствии миграции изменение численности популяции зависит от соотношения величины рождаемости и смертности.
Если величина рождаемости выше смертности, то численность популяции будет возрастать, и, наоборот, снижаться, если смертность превысит рождаемость. Таким образом, численность популяций в природных условиях постоянно меняется, поскольку меняются условия среды обитания. Амплитуда и период этих колебаний зависят от степени изменчивости окружающей среды, а также от биологических особенностей конкретного вида.
Важная роль в регуляции численности и плотности популяции принадлежит поведенческим факторам. Например, в популяциях многих грызунов повышение в крови концентрации гормона адреналина, наблюдаемое при большой плотности популяции, вызывает агрессивность, различные гормональные расстройства (у самок может происходить рассасывание эмбриона). В результате численность грызунов снижается.
Другая форма поведения — защита индивидуального участка (территориальности) — также направлена на регулирование численности. Часто почти вся территория, занимаемая популяцией, поделена на индивидуальные участки, которые обозначаются разными способами (секретом пахучих желез, царапинами на деревьях, пением самцов птиц, мочой и т. п.). Мечение и охрана участков, не допускающие размножения на них «чужих» особей, приводит к рациональному использованию территории. Избыточная часть популяции при этом не размножается или вынуждена выселяться за пределы занятого пространства.
Существует ряд других исторически сложившихся механизмов, задерживающих рост популяций и обеспечивающих тем самым их устойчивость. К ним относятся химические взаимодействия особей (например, головастики выделяют в воду вещества, которые задерживают рост других головастиков); изменения в физиологии и поведении при увеличении плотности, что приводит к проявлению инстинктов массовой миграции; распространение заболеваний (вероятность передачи инфекций возрастает с ростом плотности популяции) и др.
Таким образом, благодаря множеству механизмов размах всех суточных, сезонных и годовых изменений численности и уровня плотности популяций, как правило, меньше теоретически возможного, соответствующего реализации всего биотического потенциала. Перенаселенность всегда неблагоприятна для любого вида, так как может привести к быстрому подрыву ресурсов среды, нехватке пищи, убежищ, пространства, что неминуемо повлечет за собой общее ослабление популяций. .( http://sbio.info/page.php?id=161)