

Так что же с наследованием?
Рис. 6 Репликация ДНК (кружками обозначены метильныегруппы) |
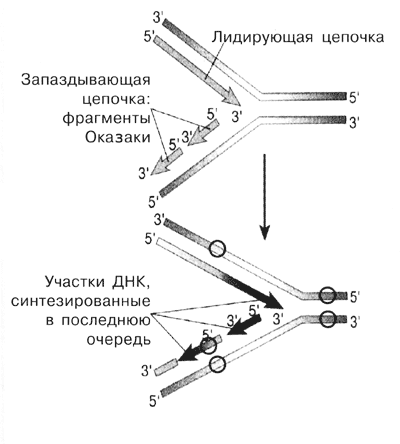
Но кроме этого, мы установили, что по степени и специфичности метилирования фрагменты Оказаки отличаются от лигированных интермедиатов репликации и от зрелой ДНК. В отличие от лигированной ДНК метилирование фрагментов Оказаки устойчиво к действию различных ингибиторов и не подавляется гормонами (ауксины у растений). Мы пришли к выводу, что в ядре имеется несколько ДНК-метилаз, которые обслуживают ДНК на разных стадиях репликации. Это полностью согласуется с современными данными.
Рис. 7 Состояние генов и их активация путем уменьшения метилирования. |
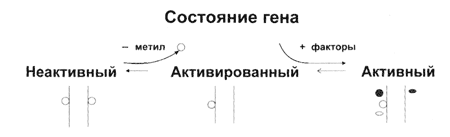
Рис. 8 Регуляция метилированием репликации ДНК в клеточном цикле. |

Таким образом, характер метилирования ДНК наследуется! И это доказано уже многими опытами. Например, установлено, что обработка растений ингибитором метилирования ДНК — 5-азацитидином —приводит к наследуемому в нескольких поколениях сильному (иногда более чем на 30%) увеличению белковости зерна. Такое и не снилось генетикам и селекционерам. Обычно гены запасных белков сильно зарепрессированы, для прорастания и начального развития растения вполне достаточно того количества запасного белка, которое синтезируется при работе генов „вполсилы“. Под действием 5-азацитидина ДНК деметилируется, и эти гены экспрессируются гораздо интенсивнее. Растению в принципе это и не нужно, зато нам — большая выгода: зерно, хлебушко наш насущный, особенно ценно для нас белком, а крахмал мы из картошки получим.
Не исключено, что неправильный, искажённый характер метилирования ДНК иногда наследуется прочно или даже навсегда. В одних случаях это может быть благом, а в других — большим злом, горем, наследственными болезнями. Однако клетка всегда стремится вернуться к исходному, правильному „портрету“ модификации её генома, и у неё существуют особые тонкие механизмы для поправки таких ударов судьбы и среды. К сожалению, мы ещё очень мало знаем об этих премудростях живого.
Как бы то ни было, сегодня доподлинно известно, что метилирование ДНК в клетке — не пустяк, оно контролирует все генетические процессы, в том числе такие, как транскрипция, репликация, рекомбинация, транспозиция генов, репарация, инактивация Х-хромосомы (половаядифференцировка). Неудивительно, что к изучению этой небольшой энзиматической модификации генома приковано внимание многих исследователей.
Здесь речь шла лишь о метилировании цитозиновых остатов в ДНК. Недавно нам удалось показать, что у эукариот геном метилируется и по адениновым остаткам. А это ещё зачем? Пока мы не знаем ответа, но предполагаем, что эта модификация также контролирует репликацию ДНК, а может быть, как у бактерий, она регулирует и экспрессию генов. Пока мы держим в руках лишь один из растительных ферментов, который метилирует адениновый остаток (внутренний А в последовательности TGATCA, предпочтительно у однотяжевых ДНК). По-видимому,этот фермент участвует в контроле за репликацией митохондриальных ДНК.