

Метаболические модели
В этом разделе мы рассмотрим и изучим особенности двух различных структурированных моделей кинетики клеточного роста, включающих некоторые стороны клеточного метаболизма. На примере этих моделей мы познакомимся с некоторыми преимуществами и недостатками, свойственными всем более детальным моделям. Эти примеры покажут нам также, что по мере включения в модель все большего числа биологических деталей она становится более специфичной для определенного организма или процесса. При создании метаболической кинетической модели определенной системы необходимо учитывать ключевые детали метаболизма, иногда известные из научных данных или биотехнологической литературы. В известном смысле мы может сказать, что, чем более детализированной становится наша модель, тем больше мы должны знать об изучаемом организме a priori. В противном случае задача выбора кинетических уравнений и значений параметров становится слишком неопределенной, причем неизвестные параметры модели нельзя определить на базе имеющихся (ограниченных) экспериментальных данных моделей, которые мы обсудим позднее, относятся к индивидуальной клетке, обычно принимаемый детерминистический принцип предполагает описание поведения усредненной клетки в большой популяции клеток.
Сначала мы изучим метаболическую модель, разработанную Бийкерком и Холлом [18], а также Памментом, Холлом и Барфордом [19] для аэробного роста почкующихся дрожжей S. cerevisiae. В основу этой модели положены перечисленные ниже допущения, большая часть которых соответствует известным данным о закономерностях клеточного цикла, метаболизма и регуляции экспрессии ферментов организма.
Принятые в модели допущения
1. Индивидуальные изолированные клетки не рассматриваются.
2. Лимитирующий рост клеток субстрат S является одновременно источником углерода и энергии; символом Е обозначен этанол.
3. Биомасса состоит из двух частей, А и В.
4. Масса А обеспечивает поглощение субстрата и снабжение клетки энергией, масса В — синтез клеток и их деление.
5. Накопление энергии и продуктов метаболизма происходит в ходе фазы Gi клеточного цикла, осуществляется массой А и описывается превращением массы А в массу В [см. приведенные ниже уравнения (7.75) и (7.76)].
6. Репликация ДНК, митоз и деление клеток (фазы S, G2 и М) происходят в течение определенного времени, осуществляются массой В и описываются превращением массы В в массу А [см. приведенное ниже уравнение (7.77)].
7. Все участвующие в процессе брожения ферменты сгруппированы и обозначены символом Ef. Аналогично сгруппированы все ферменты дыхательного пути, обозначенные Ег.
8. В каждой ферментной системе биосинтез ферментов регулируется двумя различными путями. Во-первых, ферменты продуцируются со скоростями, пропорциональными интенсивности потока метаболитов, проходящего через эту ферментную систему. Во-вторых, ферменты каждой системы продуцируются еще и в процессе адаптации клетки к изменяющимся условиям; здесь скорость их биосинтеза пропорциональна разности между имеющейся концентрацией фермента и «контрольной» величиной (eF и еR для процессов брожения и дыхания соответственно). «Контрольные» концентрации ферментов пропорциональны интенсивности потока метаболитов, который был бы достигнут при нелимитирующих концентрациях ферментов.
9. Скорости гликолиза и дыхания линейно зависят от ef/eF и от er/eR соответственно.
10. Гликолитические ферменты продуцируются в результате работы дыхательного пути (таким путем образуются ферменты для глюконеогенеза, биосинтеза и утилизации внутриклеточных резервных углеводов).
В таком виде клеточный рост можно описать с помощью следующих стехиометрических уравнений [здесь и далее индексами W обозначены переменные, выраженные в единицах массы; в других случаях подразумевается число молей или единиц активности (для ферментов)]:

Применение этой кинетической модели к экспериментам по росту непрерывной культуры в стационарном состоянии потребовало незначительного изменения (от 1,79 до 1,50 в периодическом и непрерывном процессах соответственно) только одного параметра а3 — экономического коэффициента роста за счет дыхания. Причиной его изменения отчасти послужила известная физиология этого организма, в частности необходимость глюконеогенеза при росте периодической культуры на этаноле и отсутствие такой необходимости при росте проточной культуры на глюкозе (в ПРПП).

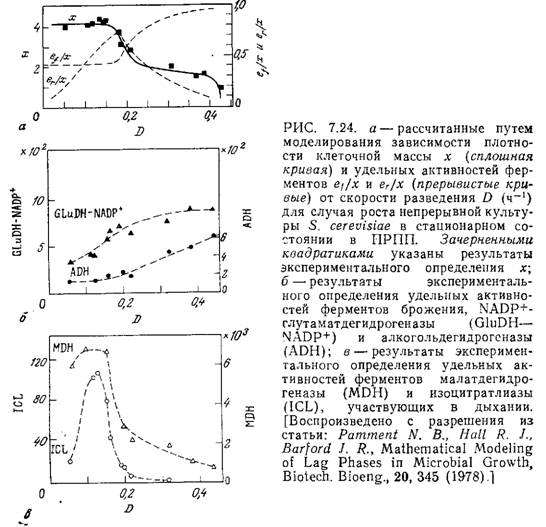
На рис. 7.24, а изображены графики зависимости плотности клеточной массы х(х = а + b) и удельных активностей ферментов брожения (ef/x) и дыхания (еr/х) от скорости разведения для случая роста непрерывной культуры 5. cerevisiae в стационарном состоянии; эти зависимости вычислены с помощью параметров, перечисленных в табл. 7.3. Зачерненными квадратиками на этом рисунке обозначены результаты экспериментального определения х, хорошо согласующиеся с расчетными данными. Обратите внимание на то, что здесь зависимость х от D как для экспериментальных, так и для расчетных данных резко отличается от аналогичной упрощенной зависимости, полученной на базе неструктурированной модели Моно (рис. 7.6) На рис. 7.24, б приведены результаты экспериментального определения зависимости удельной активности двух ферментов, участвующих в метаболизме брожения (NАDР+-глутаматдегидро-геназы и алкогольдегидрогеназы) от скорости разведения в ПРПП. Эти зависимости качественно согласуются с расчетными данными для еf\х модели. На кривой зависимости удельной активности двух участвующих в дыхании ферментов (малатдегид-рогеназы и изоцитратлиазы) от скорости разведения экспериментально обнаружен максимум (рис. 7.24, в); модель предсказывает наличие максимума в зависимости еr/х от D.
Эти результаты, как и ряд других, приведенных в работе [18], убедительно демонстрируют некоторые из преимуществ, свойственных более структурированному описанию роста популяции клеток. Рассматриваемая модель успешно применялась для описания периодического роста 5. cerevisiae с добавлением субстрата (см. разд. 9.1.1); она также хорошо отражает лаг-фазу в экспериментальных периодических процессах.
В качестве завершающего примера рассмотрим разработанную Шулером и сотрудниками модель индивидуальной клетки Е. coli [24]; из всех предложенных до настоящего времени моделей роста микроорганизмов она наиболее удачна и высокоструктирована. На рис. 7.25 схематично изображены учитываемые в этой модели метаболиты, биополимеры и реакции, осуществляющиеся, как предполагается, в клетке Е. coli В/г А. Прерывистыми линиями здесь обозначены информационные потоки, регулирующие кинетику реакций. В модель включены реакции образования клеточной оболочки, поэтому продолжительность клеточного цикла является величиной, предсказываемой моделью, а не исходным параметром, как в большинстве других структурированных моделей. Поскольку эта модель разрабатывалась специально для описания роста клеток в условиях ограниченного количества источника углерода или азота, в ней учтены многие структурные элементы клетки, участвующие в транспорте и ассимиляции этих веществ. Приведенные на рис. 7.25 реакции учитывают также синтез и утилизацию АТР.
На рис. 7.25 не показаны важные детали модели, связанные с регуляцией инициации синтеза ДНК. В течение очень короткого периода («взрывной синтез») синтезируется белок-реп-рессор RP; он нейтрализуется белком-антирепрессором ARP, который синтезируется со скоростью, пропорциональной скорости построения клеточной оболочки. При достаточно низкой концентрации RP начинается транскрипция, которая приводит к короткому участку РНК, необходимому для инициации репликации.
К сожалению, объем нашей книги не позволяет описать эту модель достаточно подробно; дополнительные данные читатель может найти в работе [20].
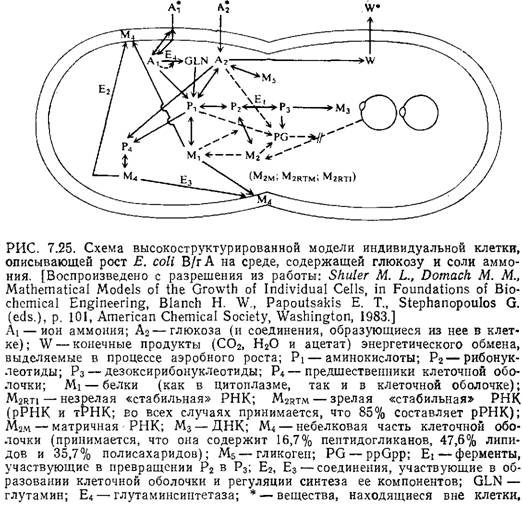
Эта модель включает около 100 стехиометрических и кинетических параметров; почти все эти параметры могут быть определены на основе опубликованных результатов изучения биохимии Е coli. Насколько велика информация, вложенная в эту модель, настолько же велики нее возможности.
Модель достаточно точно описывает время инициации репликации хромосомы и другие важнейшие особенности клеточного цикла в широком диапазоне скоростей роста. На рис. 7.26 приведены графики зависимости в двойных обратных координатах удельной скорости роста отдельной клетки от концентрации лимитирующего рост питательного вещества (в данном случае иона аммония), полученные экспериментально (точки) и расчетным путем на базе описанной модели (сплошная линия). Обратите внимание на то, что здесь и результаты моделирования, и экспериментальные данные свидетельствуют о существовании нескольких механизмов утилизации иона аммония; это проявляется в изменении наклона при очень больших концентрациях питательного вещества (малых величинах l/s). Вычисленные путем моделирования закономерности изменения скорости клеточного роста, содержания в клетке гликогена и размера клетки (рис. 7.27) хорошо согласуются с экспериментальными результатами.
Р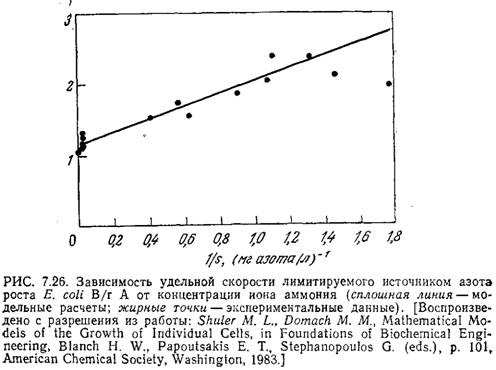 ассмотренная
нами ранее модель Уильямса предсказывает
увеличение размеров клеток при высоких
скоростях роста; в отличие от нее
описываемая высокоструктурированная
модель, учитывающая гораздо большее
число биохимических особенностей и
деталей метаболизма клетки, показывает,
что размер клеток зависит не только от
скорости их роста, но и от того, какое
питательное вещество лимитирует скорость
роста.
ассмотренная
нами ранее модель Уильямса предсказывает
увеличение размеров клеток при высоких
скоростях роста; в отличие от нее
описываемая высокоструктурированная
модель, учитывающая гораздо большее
число биохимических особенностей и
деталей метаболизма клетки, показывает,
что размер клеток зависит не только от
скорости их роста, но и от того, какое
питательное вещество лимитирует скорость
роста.