

Определение
Закон независимого наследования (третий закон Менделя) — при скрещивании двух дигетерозиготных особей, отличающихся друг от друга по двум (и более) парам альтернативных признаков, гены и соответствующие им признаки наследуются независимо друг от друга и комбинируются во всех возможных сочетаниях (как и при моногибридном скрещивании).
Когда скрещивались растения, отличающиеся по нескольким признакам, таким как белые и пурпурные цветы и желтые или зелёные горошины, наследование каждого из признаков следовало первым двум законам и в потомстве они комбинировались таким образом, как будто их наследование происходило независимо друг от друга. Первое поколение после скрещивания обладало доминантным фенотипом по всем признакам. Во втором поколении наблюдалось расщепление фенотипов по формуле 9:3:3:1, то есть 9:16 были с пурпурными цветами и желтыми горошинами, 3:16 с белыми цветами и желтыми горошинами, 3:16 с пурпурными цветами и зелёными горошинами, 1:16 с белыми цветами и зелёными горошинами.
Объяснение
Менделю попались признаки, гены которых находились в разных парах гомологичных хромосом гороха. При мейозе гомологичные хромосомы разных пар комбинируются в гаметах случайным образом. Если в гамету попала отцовская хромосома первой пары, то с равной вероятностью в эту гамету может попасть как отцовская, так и материнская хромосома второй пары. Поэтому признаки, гены которых находятся в разных парах гомологичных хромосом, комбинируются независимо друг от друга. (Впоследствии выяснилось, что из исследованных Менделем семи пар признаков у гороха, у которого диплоидное число хромосом 2n=14, гены, отвечающие за одну из пар признаков, находились в одной и той же хромосоме. Однако Мендель не обнаружил нарушения закона независимого наследования, так как сцепления между этими генами не наблюдалось из-за большого расстояния между ними).
Статистический характер законов наследования. Условия выполнения законов Менделя Судьба открытия Менделя. Переоткрытие законов Менделя.
Наука развивается закономерно. Когда в ней накапливается достаточное количество фактов, идеи некоторых опытов, обобщений или законов начинают "носиться в воздухе" и нередко практически одновременно открываются несколькими учеными в разных странах. Недаром так много законов и правил носит "двойные имена" (закон Био-Савара, закон Джоуля- Ленца, закон Бойля-Мариотта и т.д.). Два физика одновременно открыли ультрафиолетовые лучи, два химика в один и тот же год открыли кислород и т.д.
Яркий пример закономерности развития науки дает и возникновение генетики. Годом ее рождения считается 1900-й, когда три ботаника, работавшие в разных странах, одновременно пришли в своих исследованиях к выводам, сходным с выводами Менделя, а при подготовке своих результатов к публикации наткнулись на его работу. Эти ботаники - Г.де Фриз из Голландии, который работал с маком; К.Корренс из Германии, работавший с кукурузой; Э.Чермак из Австрии, который, как и Мендель, работал с горохом. Независимо от них английский генетик Бэтсон в 1898 г. вел опыты по скрещиванию кур и пришел к тем же выводам. Работа Бэтсона вышла из печати в 1902 г. В том же году появилось сообщение французского зоолога Гено о скрещивании серых мышей с альбиносами, в результате которого в первом поколении были получены только серые мыши, а во втором - 198 серых особей и 72 альбиноса. Таким образом, была доказана применимость законов Менделя к животным. После переоткрытия законов Менделя их подтвердили на огромном количестве разнообразных признаков многих животных и растений. Заметим, что и сам Мендель ставил опыты не только на горохе. Он проверил справедливость своих выводов на фасоли, левкое, кукурузе и ночной красавице.
Ряд ученых просто повторили опыты Менделя на очень большом материале и подтвердили правильность его выводов. Так, например, были повторены опыты на горохе с расщеплением по отдельным признакам. Приведем результат только для расщепления по желтой и зеленой окраске семян. Бэтсон в 1905 г. на 16000 семян получил 75,3% доминантных (желтых) и 24,7% рецессивных (зеленых). Дэрбишер в 1909 г., рассмотрев 145000 семян, получил расщепление в отношении 75,1% к 24,9%. Вообще, начиная с 1900 г., опыты по гибридизации получили очень большой размах. Прежде всего, были изучены признаки, относящиеся к окраске и форме разных организмов, так как эти признаки легко различимы невооруженным глазом.
Приведем несколько примеров первых работ по изучению расщепления признаков. Было показано выполнение законов Менделя для цвета гусениц и цвета коконов шелкопряда ( рис. 92 , А). Установили, что отсутствие полос на раковине виноградной улитки доминирует над их присутствием, что длинная шерсть у ангорских кроликов - рецессивный признак по отношению к короткой шерсти, а отсутствие рогов у крупного рогатого скота - доминантный признак ( рис. 92 , Б) и т.д.
Было обнаружено, что законы Менделя применимы не только к морфологическим, но и к физиологическим признакам. Так, Корренс (1904) показал на белене, что двухлетность доминирует над однолетностыо. Н.И.Вавилов показал, что невосприимчивость растений к заболеваниям, вызываемым грибками, наследуется по законам Менделя.
Позднее сходные результаты были получены и для животных. Например, было показано, что мыши чистой линии, выведенные в Принстонском университете, невосприимчивы к ряду вирусных заболеваний, поражающих нервную систему (к вирусу желтой лихорадки , вирусу энцефалита и др.). Мыши другой линии при заражении этими вирусами погибали в 100% случаев. Гибриды этих двух линий все были устойчивы к болезни. А из гибридов второго поколения 1/4 погибала, а 3/4 были невосприимчивы к заболеваниям.
Как ясно из приведенных примеров, уже при возникновении генетики началось изучение генетических особенностей ряда хозяйственно важных объектов и признаков, имеющих большое практическое значение.
Закон Менделя и наследование у человека. У человека давно известен ряд внешних признаков, которые наследуются по Менделю. Например, курчавые или волнистые волосы доминируют над прямыми. Темный цвет глаз доминирует над голубым. Такие признаки давно использовались для определения родства (или как доказательство невозможности родства). Так, например, в романе Агаты Кристи "Рождество Эркюля Пуаро" знаменитый сыщик по цвету глаз определяет, что одна из героинь в действительности не является внучкой хозяина поместья. Пуаро говорит ей: "Мадемуазель, если бы вы изучали законы Менделя, вам было бы известно, что в семье, где у обоих родителей голубые глаза, не бывает кареглазых детей". Кстати, тут Пуаро ошибся. Бывает, что у родителей с голубыми глазами и очень небольшим содержанием темного пигмента в радужной оболочке, рождаются кареглазые дети. Один из дефектов зрения, так называемая врожденная куриная слепота , - сильное снижение зрения в сумерках - наследуется как доминантный признак. В одной французской семье этот дефект был прослежен на протяжении 10 поколений.
Таким образом, в соответствии с законом Менделя наследуются самые разные признаки - в том числе и такие, которые имеют важное значение для сельского хозяйства и медицины.
Вопрос № 25.
ЗАКОНОМЕРНОСТИ НАСЛЕДОВАНИЯ.
а) Гибридологический анализ – основной метод генетики.
Основу генетического анализа составляет гибридологический анализ, основанный на анализе наследования признаков при скрещиваниях.
Гибридологический анализ, основы которого разработал основатель современной генетики Г. Мендель, основан на следующих принципах.
1. Использование в качестве исходных особей (родителей), форм, не дающих расщепления при скрещивании, т.е. константных форм.
2. Анализ наследования отдельных пар альтернативных признаков, то есть признаков, представленных двумя взаимоисключающими вариантами.
3. Количественный учет форм, выщепляющихся в ходе последовательных скрещиваний и использование математических методов при обработке результатов.
4. Индивидуальный анализ потомства от каждой родительской особи.
5. На основании результатов скрещивания составляется и анализируется схема скрещиваний.
Однако в некоторых случаях метод прямого гибридологического анализа оказывается неприменим. Например, при изучении наследования признаков у человека необходимо учитывать ряд обстоятельств: невозможность планирования скрещиваний, низкая плодовитость, длительный период полового созревания. Поэтому кроме гибридологического анализа, в генетике используется множество других методов.
Гибридологический анализ, способ изучения наследственных свойств организма путём скрещивания (гибридизации) его с родственной формой и последующим анализом признаков потомства. Г. а. впервые применил Г. Мендель (1865) для изучения механизма передачи наследственных задатков (генов) от родителей потомкам и для изучения взаимодействия генов у одного и того же организма. В основе Г. а. лежит способность к рекомбинации, т. е. перераспределению генов при образовании гамет, что приводит к возникновению новых сочетаний генов. По этим сочетаниям, которые проявляются в потомстве гибридной особи с определённой частотой, можно судить о генотипе родительской формы, а по генотипу родительской формы можно предсказывать генотип потомства. Так, генотип особи, гибридной по паре аллелей, одна из которых - доминантная А, другая - рецессивная а, можно представить как Аа. Внешне, т. е. фенотипически, такая форма (гетерозигота) не отличается от формы с генотипом АА (гомозигота). Гибрид (Аа) формирует гаметы двух типов, каждый из которых несёт аллель А или аллель а. Т. о., гаметы никогда не бывают гибридными. С помощью различных видов скрещивания можно выявить, сколько типов гамет по данному гену формирует организм, и определить его генотип. Если у анализируемой формы (Аа) возможно самооплодотворение (что часто встречается у растений), схематично это будет выглядеть так: ♂ (А+а) ´ ♀ (А+а)(АА + Аа + Аа + аа. При этом в потомстве с определённой частотой появляется новая форма - аа.
Если самооплодотворения нет, генотип исходной формы выявляют, скрещивая в разных комбинациях её потомков ("брат ´ сестра") и анализируя "внучатое" поколение. Др. способ выявления гибридного состояния - анализирующее скрещивание: скрещивание предполагаемого гибрида с рецессивной родительской формой. Г. а. играет важную роль в селекционной практике и племенном деле, т.к. позволяет судить о тождестве фенотипа и генотипа. Здесь Г. а. находит применение в форме "анализа производителей по потомству" с целью выявления у производителей скрытых нежелательных генов. Г. а. применяется также при составлении хромосомных карт. Знание генного состава хромосомы позволяет путём специальных скрещиваний вводить в геном определённую хромосому или группу генов и создавать формы с нужным генотипом. Этот метод широко применяется в растениеводстве. Г. а. пользуются при изучении взаимодействия генов в первом гибридном поколении (тесты на комплементацию). Г. а. является главным методом генетического анализа.
б) ?Условия моделирования признаков?.
в) Неполное доминирование (пример).
Неполное доминирование. Доминантный ген в гетерозиготном состоянии не всегда полностью подавляет рецессивный ген. В ряде случаев гибрид fi не воспроизводит полностью ни одного из родительских признаков и признак носит промежуточный характер с большим или меньшим уклонением к доминантному или рецессивному состоянию. Но все особи этого поколения единообразны по данному признаку. Так, при скрещивании ночной красавицы с красной окраской цветков (АА) с растением, имеющим белые цветки (аа), в fi образуется промежуточная розовая окраска цветка (Аа). При неполном доминировании в потомстве гибридов (Fi) расщепление по генотипу и фенотипу совпадает (1:2:1).
Неполное доминирование — широко распространенное явление. Оно обнаружено при изучении наследования окраски цветка у львиного зева, окраски шерсти у крупного рогатого скота и овец, биохимических признаков у человека и т. д. Промежуточные признаки, возникающие вследствие неполного доминирования, нередко представляют эстетическую или материальную ценность для человека. Возникает вопрос: можно ли вывести путем отбора, например, сорт ночной красавицы с розовой окраской цветков? Очевидно, нет, потому что этот признак развивается только у гетерозигот и при скрещивании их между собой всегда происходит расщепление:
В своих опытах Мендель имел дело с примерами полного доминирования, поэтому гетерозиготные особи в его опытах оказались не отличимы от доминантных гомозигот. Но в природе наряду с полным доминированием часто бывает неполное доминирование. Так, у душистого горошка известны две расы: с красными и белыми цветами. Гибриды, полученные от скрещивания этих рас, имеют промежуточную розовую окраску. Во втором поколении расщепление по фенотипу соответствует расщеплению и по генотипу, т. е. происходит в отношении 1 красный : 2 розовых : 1 белый (см. таблицу I на вклейке, рис. Б).
У андалузских кур бывает черная и белая окраска перьев, а их гибриды имеют голубую окраску. При скрещивании крупного рогатого скота красной масти с белыми животными гибриды получаются чалой масти, т. е. у них часть волос белых, а часть красных, перемешанных более или менее равномерно.
Свойством неполного доминировании обладает ряд генов, вызывающих наследственные аномалии и болезни человека. Так, например, наследуется серповидноклеточная анемия (о ней подробнее будет сказано ниже) и атаксия Фредрейха, характеризуемая прогрессирующей потерей координации произвольных движений.
г) Анализирующее скрещивание (пример).
Анализирующее скрещивание — скрещивание гибридной особи с особью, гомозиготной по рецессивным аллелям, то есть "анализатором". Смысл анализирующего скрещивания заключается в том, что потомки от анализирующего скрещивания обязательно несут один рецессивный аллель от "анализатора", на фоне которого должны проявиться аллели, полученные от анализируемого организма. Для анализирующего скрещивания (исключая случаи взаимодействия генов) характерно совпадение расщепления по фенотипу с расщеплением по генотипу среди потомков. Таким образом, анализирующее скрещивание позволяет определить генотип и соотношение гамет разного типа, образуемых анализируемой особью.
д) Сверхдоминирование (пример).
Сверхдоминирование — это явление преимущества класса гетерозигот по сравнению с возможными, для данного гена и аллелей, классами гомозигот.
Фенотипически, как правило, в случае сверхдоминирования гетерозиготы не обладают особыми внешними признаками. Преимущество связано с биохимическими особенностями.
Один из характерных примеров сверхдоминирвания является повышенная частота аллеля гена серповидноклеточной анемии в популяциях человека, живущих в условиях высокой вероятности заражения малярией. Мутантный аллель защищает организм от заболевания малярией. Гомозиготы по нормальному аллелю могут заболеть малярией и погибнуть, гомозиготы по мутантному аллелю - с высокой вероятностью гибнут от анемии. Гетрозиготы по этому гену не болеют серповидновлеточной анемией и устойчивы к малярии.
Преимущество гетерозигот так же показано по многим генам и у многих организмов. Для Drosophila melanogaster показаны эффекты сверхдоминирования по гену алкогольдегидрогеназы в лабораторных популяциях].
В ряде случаев аллель гена, с которым связано сверхдоминирование является рецессивно летальным, и поддерживается в популяции за счёт преимущества гетерозигот. К таким случаям относится например система летальных аллелей гена lethal giant larvae. Гетерозиготы, имеющие нормальный и мутантный вариант этого гена, в ряде случаев, характеризуются повышенной жизнеспособностью.
Вопрос № 26.
ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ (Т. МОРГАН)
а) История создания хромосомной теории.
В 1902-1903 гг. американский цитолог У. Сеттон и немецкий цитолог и эмбриолог Т. Бовери независимо друг от друга выявили параллелизм в поведении генов и хромосом в ходе формирования гамет и оплодотворения. Эти наблюдения послужили основой для предположения, что гены расположены в хромосомах. Однако экспериментальное доказательство локализации конкретных генов в конкретных хромосомах было получено только в 1910 г. американским генетиком Т. Морганом, который в последующие годы (1911—1926) обосновал хромосомную теорию наследственности. Согласно этой теории, передача наследственной информации связана с хромосомами, в которых линейно, в определенной последовательности, локализованы гены. Таким образом, именно хромосомы представляют собой материальную основу наследственности.
Формированию хромосомной теории способствовали данные, полученные при изучении генетики пола, когда были установлены различия в наборе хромосом у организмов различных полов.
б) Линейное расположение генов в хромосомах.
?Существование кроссинговера побудило Моргана разработать в 1911-1914 гг. принцип построения генетических карт хромосом. В основу этого принципа положено представление о расположении генов по длине хромосомы в линейном порядке. За единицу расстояния между двумя генами условились принимать 1 % перекреста между ними.
Допустим, что к одной группе сцепления относятся гены А и В. Между ними обнаружен перекрест в 10 %. Следовательно, гены А и В находятся на расстоянии 10 единиц. Допустим далее, что к этой же группе сцепления относится ген С. Чтобы узнать его место в хромосоме, необходимо выяснить, какой процент перекреста он дает с обоими из двух уже известных генов. Например, если с А он дает 3 % перекреста, то можно предположить, что ген С находится либо между А и В, либо с противоположной стороны, то есть А расположен между В и С. Если между В и С окажется перекрест 7 %, то на хромосоме их следует расположить в таком порядке, как на верхней схеме. Если между В и С перекрест составит 13 %, то расположение генов будет как на нижней схеме.
в) Хромосомы как группы сцепления генов.
Гены, локализованные в одной хромосоме, называются группой сцепления.
Сцепление генов, совместная передача двух или более генов от родителей потомкам. Объясняется тем, что эти гены лежат в одной хромосоме, то есть принадлежат одной группе сцепления и поэтому не могут случайно перекомбинироваться в мейозе, как это бывает при наследовании генов, лежащих в разных хромосомах. Сцепление генов было открыто в 1906 английскими генетиками У. Бэтсоном и Р. Пеннетом, обнаружившими в опытах по скрещиванию растений у некоторых генов тенденцию передаваться совместно и тем самым нарушать закон независимого комбинирования признаков. Правильное объяснение этому дали Т. Морган и сотрудники, обнаружившие аналогичное явление при изучении наследования признаков у дрозофилы. Мерой Сцепление генов служит частота образования гетерозиготой по этим генам кроссоверных гамет или спор, в которых гены находятся не в исходных, а в новых сочетаниях благодаря обмену частями несущих их гомологичных хромосом путём кроссинговера. У некоторых бактерий др. мерой Сцепление генов служит частота совместной передачи по наследству разных генов при конъюгации, генетической трансформации и трансдукции. Сила Сцепление генов может быть различной у разных полов (обычно она больше у гетерогаметного пола, см. Половые хромосомы) или даже Сцепление генов может быть полным (отсутствие кроссинговера) у одного из полов (например, у самцов дрозофилы или у самок тутового шелкопряда). Кроме того, сила Сцепление генов может варьировать в зависимости от возраста родителей, температуры, наличия хромосомных перестроек и др. факторов, а также от присутствия особых мутантных генов, специфически влияющих на силу Сцепления генов.
г) Генетические и цитологические карты хромосом.
Генетические карты хромосом, схемы относительного расположения сцепленных между собой наследственных факторов - генов. Генетические карты хромосом отображают реально существующий линейный порядок размещения генов в хромосомах и важны как в теоретических исследованиях, так и при проведении селекционной работы, т.к. позволяют сознательно подбирать пары признаков при скрещиваниях, а также предсказывать особенности наследования и проявления различных признаков у изучаемых организмов. Имея Генетические карты хромосом, можно по наследованию «сигнального» гена, тесно сцепленного с изучаемым, контролировать передачу потомству генов, обусловливающих развитие трудно анализируемых признаков; например, ген, определяющий эндосперм у кукурузы и находящийся в 9-й хромосоме, сцеплен с геном, определяющим пониженную жизнеспособность растения. Генетические карты хромосом составляют для каждой пары гомологичных хромосом. Группы сцепления нумеруют последовательно, по мере их обнаружения. Кроме номера группы сцепления, указывают полные или сокращённые названия мутантных генов, их расстояния в морганидах от одного из концов хромосомы, принятого за нулевую точку, а также место центромеры. Составить Генетические карты хромосом можно только для объектов, у которых изучено большое число мутантных генов.
Цитологические карты хромосом, схематическое изображение хромосом с указанием мест фактического размещения отдельных генов, полученное с помощью цитологических методов. Цитологические карты хромосом составляют для организмов, для которых обычно уже имеются генетические карты хромосом. Каждое место расположения гена (локус) на генетической карте организма, установленное на основе частоты перекреста участков хромосом (кроссинговера), на Цитологические карты хромосом привязано к определённому, реально существующему участку хромосомы, что служит одним из основных доказательств хромосомной теории наследственности. Для построения Цитологических карт хромосом используют данные анализа хромосомных перестроек (вставки, делеции и др.) и, сопоставляя изменения морфологических признаков хромосом при этих перестройках с изменениями генетических свойств организма, устанавливают место того или иного гена в хромосоме.
д) Основные положения хромосомной теории.
Анализ явлений сцепленного наследования, кроссинговера, сравнение генетической и цитологической карт позволяют сформулировать основные положения хромосомной теории наследственности:
Гены локализованы в хромосомах. При этом различные хромосомы содержат неодинаковое число генов. Кроме того, набор генов каждой из негомологичных хромосом уникален.
Аллельные гены занимают одинаковые локусы в гомологичных хромосомах.
Гены расположены в хромосоме в линейной последовательности.
Гены одной хромосомы образуют группу сцепления, то есть наследуются преимущественно сцепленно (совместно), благодаря чему происходит сцепленное наследование некоторых признаков. Число групп сцепления равно гаплоидному числу хромосом данного вида (у гомогаметного пола) или больше на 1 (у гетерогаметного пола).
Сцепление нарушается в результате кроссинговера, частота которого прямо пропорциональна расстоянию между генами в хромосоме (поэтому сила сцепления находится в обратной зависимости от расстояния между генами).
Каждый биологический вид характеризуется определенным набором хромосом — кариотипом.
Вопрос № 27.
МОЛЕКУЛЯРНЫЕ ЗАКОНЫ НАСЛЕДСТВЕННОСТИ.
а) История открытия нуклеиновых кислот.
В 1928 английский бактериолог Ф. Гриффит обнаружил, что убитые патогенные пневмококки могут изменять генетические свойства живых непатогенных пневмококков, превращая последние в патогенные. В 1945 микробиолог О. Эвери из Рокфеллеровского института в Нью-Йорке сделал важное открытие: он показал, что способность к генетической трансформации обусловлена переносом ДНК из одной клетки в другую, а следовательно, генетический материал представляет собой ДНК. В 1940-1950 Дж. Бидл и Э. Тейтум из Станфордского университета (шт. Калифорния) обнаружили, что синтез белков, в частности ферментов, контролируется специфическими генами. В 1942 Т. Касперсон в Швеции и Ж. Браше в Бельгии открыли, что нуклеиновых кислот особенно много в клетках, активно синтезирующих белки. Все эти данные наводили на мысль, что генетический материал - это нуклеиновая кислота и что она как-то участвует в синтезе белков. Однако в то время многие полагали, что молекулы нуклеиновых кислот, несмотря на их большую длину, имеют слишком простую периодически повторяющуюся структуру, чтобы нести достаточно информации и служить генетическим материалом. Но в конце 1940-х годов Э. Чаргафф в США и Дж. Уайатт в Канаде, используя метод распределительной хроматографии на бумаге, показали, что структура ДНК не столь проста и эта молекула может служить носителем генетической информации.
Структура ДНК была установлена в 1953 М. Уилкинсом, Дж. Уотсоном и Ф. Криком в Англии. Это фундаментальное открытие позволило понять, как происходит удвоение (репликация) нуклеиновых кислот. Вскоре после этого американские исследователи А. Даунс и Дж. Гамов предположили, что структура белков каким-то образом закодирована в нуклеиновых кислотах, а к 1965 эта гипотеза была подтверждена многими исследователями: Ф. Криком в Англии, М. Ниренбергом и С. Очоа в США, Х. Кораной в Индии. Все эти открытия, результат столетнего изучения нуклеиновых кислот, произвели подлинную революцию в биологии. Они позволили объяснить феномен жизни в рамках взаимодействия между атомами и молекулами.
б) Доказательства роли ДНК как хранителя наследственной информации.
1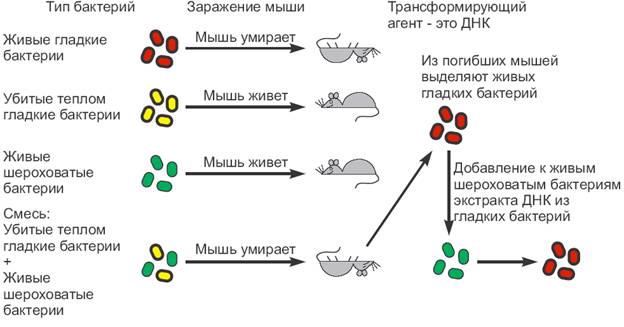 . 1928г.
Опыты Фредерика Гриффита
Известно,
что
. 1928г.
Опыты Фредерика Гриффита
Известно,
что
бактерия Pneutnococcus pneumoniaeимеет
несколько форм. Вирулентность бактерии
определяется наличием мукополисахаридной
капсулы, расположенной па поверхности
клетки. Эта капсула защищает бактерию
от воздействий со стороны организма-хозяина.
В результате, размножившиеся бактерии
убивают зараженное животное. Бактерии
этого штамма (S-штамм) образуют гладкие
колонии. Авирулентные формы бактерий
не имеют защитной капсулы и образуют
шероховатые колонии (R-штамм). Микробиолог
Фредерик Гриффитс в 1928 году инъецировал
мышам живого пневмококка R-штамма вместе
с S-штаммом, убитым высокой температурой
(65°С). Спустя некоторое время ему удалось
выделить из заражённых мышей живых
пневмококков, обладающих капсулой.
Таким образом, оказалось, что свойство
убитого пневмококка - способность
образовывать капсулу - перешло к живой
бактерии, т.е. произошла трансформация.
Поскольку признак наличия капсулы
является наследственным, то следовало
предположить, что какая-то часть
наследственного вещества от бактерий
штамма S перешла к клеткам штамма R.
В
1944 году О.Т. Эвери, К.М. Маклеод и М.
Маккарти показали, что такое же превращение
типов пневмококков может происходить
в пробирке, т.е. in vitro. Эти
исследователи установили существование
особой субстанции -"трансформирующего
принципа", -экстракта из клеток штамма
S, обогащенного ДНK. Как далее в ыяснилось,
ДНK, выделенная из клеток S-штамма
добавленная в культуру R-штамма,
трансформировала часть клеток в S-форму,
Клетки стойко передавали это свойство
при дальнейшем размножении. Обработка
"трансформирующего фактора"
ДНК-азой, ферментом разрушающим ДНK,
блокирована трансформацию. Эти данные
впервые показали, что именно ДНК, а не
белок, как полагали до тех пор, является
наследственным материалом.
2.
1952г. Эксперимент Альфреда Херши и Марты
Чейз.
Как
известно, фаг Т2 является вирусом,
инфицирующим бактерию E. coli. фаговые
частицы абсорбируются на наружной
поверхности клетки, их материал проникает
внутрь и примерно через 20 минут бактерия
лизируется, освобождая большое количество
фаговых частиц - потомков. В 1952 году
Альфред Херши и Марта Чейз инфицировали
бактерии фагами Т2, которые были мечены
радиоактивными соединениями: ДНК - с
помощью 32P. Белковая часть фага - 35S. После
инфекции бактерии фагами, с помощью
центрифугирования удалось выделить
две фракции: пустые белковые оболочки
фага и бактерии, инфицированных фаговой
ДНК. Оказалось, что 80% метки 35S осталась
в пустых фаговых оболочках, а 70% метки
32P - в инфицированных бактериях.
Фаги-потомки получили только около 1%
исходного белка, меченного 35S, однако
они же обнаружили около 30% метки
32P.
Результаты
этого эксперимента прямо показали, что
ДНК родительских фагов проникает в
бактерии и затем становиться составляющей
развившихся новых фагов частиц.
3.
1957г. Опыты Френкеля - Конрата
Френкель-Конрат
работал с вирусом табачной мозаики
(ВТМ). В этом вирусе содержится РНК, а не
ДНК. Было известно, что разные штаммы
вируса вызывают разную картину поражения
листьев табака. После смены белковой
оболочки "переодетые" вирусы
вызывали картину поражения, характерную
для того штамма, чья РНК была покрыта
чужим белком.
Следовательно,
не только ДНК, но и РНК может служить
носителем генетической информации.
На
сегодняшний день существуют сотни тысяч
доказательств генетической роли
нуклеиновых кислот. Приведенные три
являются классическими.
ыяснилось,
ДНK, выделенная из клеток S-штамма
добавленная в культуру R-штамма,
трансформировала часть клеток в S-форму,
Клетки стойко передавали это свойство
при дальнейшем размножении. Обработка
"трансформирующего фактора"
ДНК-азой, ферментом разрушающим ДНK,
блокирована трансформацию. Эти данные
впервые показали, что именно ДНК, а не
белок, как полагали до тех пор, является
наследственным материалом.
2.
1952г. Эксперимент Альфреда Херши и Марты
Чейз.
Как
известно, фаг Т2 является вирусом,
инфицирующим бактерию E. coli. фаговые
частицы абсорбируются на наружной
поверхности клетки, их материал проникает
внутрь и примерно через 20 минут бактерия
лизируется, освобождая большое количество
фаговых частиц - потомков. В 1952 году
Альфред Херши и Марта Чейз инфицировали
бактерии фагами Т2, которые были мечены
радиоактивными соединениями: ДНК - с
помощью 32P. Белковая часть фага - 35S. После
инфекции бактерии фагами, с помощью
центрифугирования удалось выделить
две фракции: пустые белковые оболочки
фага и бактерии, инфицированных фаговой
ДНК. Оказалось, что 80% метки 35S осталась
в пустых фаговых оболочках, а 70% метки
32P - в инфицированных бактериях.
Фаги-потомки получили только около 1%
исходного белка, меченного 35S, однако
они же обнаружили около 30% метки
32P.
Результаты
этого эксперимента прямо показали, что
ДНК родительских фагов проникает в
бактерии и затем становиться составляющей
развившихся новых фагов частиц.
3.
1957г. Опыты Френкеля - Конрата
Френкель-Конрат
работал с вирусом табачной мозаики
(ВТМ). В этом вирусе содержится РНК, а не
ДНК. Было известно, что разные штаммы
вируса вызывают разную картину поражения
листьев табака. После смены белковой
оболочки "переодетые" вирусы
вызывали картину поражения, характерную
для того штамма, чья РНК была покрыта
чужим белком.
Следовательно,
не только ДНК, но и РНК может служить
носителем генетической информации.
На
сегодняшний день существуют сотни тысяч
доказательств генетической роли
нуклеиновых кислот. Приведенные три
являются классическими.
в) Химический состав и строение нуклеиновых кислот.
Полимерные формы нуклеиновых кислот называют полинуклеотидами. Цепочки из нуклеотидов соединяются через остаток фосфорной кислоты (фосфодиэфирная связь). Поскольку в нуклеотидах существует только два типа гетероциклических молекул, рибоза и дезоксирибоза, то и имеется лишь два вида нуклеиновых кислот — дезоксирибонуклеиновая (ДНК) и рибонуклеиновая (РНК).
Мономерные формы также встречаются в клетках и играют важную роль в процессах передачи сигналов или запасании энергии. Наиболее известный мономер РНК — АТФ, аденозинтрифосфорная кислота, важнейший аккумулятор энергии в клетке.
ДНК — Дезоксирибонуклеиновая кислота. Сахар — дезоксирибоза, азотистые основания: пуриновые — гуанин (G), аденин (A), пиримидиновые — тимин (T) ицитозин (C). ДНК часто состоит из двух полинуклеотидных цепей, направленных антипараллельно.
РНК — Рибонуклеиновая кислота. Сахар — рибоза, азотистые основания: пуриновые — гуанин (G), аденин (A), пиримидиновые урацил (U) и цитозин (C). Структура полинуклеотидной цепочки аналогична таковой в ДНК. Из-за особенностей рибозы молекулы РНК часто имеют различные вторичные и третичные структуры, образуя комплементарные участки между разными цепями.
г) Создание модели ДНК и её обоснование (Уотсон, Крик).
В 1950 г. английский физик М.Уилкинс получил рентгенограмму кристаллических волокон ДНК. Она показала, что молекула ДНК имеет определенную структуру, расшифровка которой помогла бы понять механизм функционирования ДНК. Рентгенограммы, полученные не на кристаллических волокнах ДНК, а на менее упорядоченных агрегатах, которые образуются при более высокой влажности, позволили Розалинд Франклин , коллеге М. Уилкинса, увидеть четкий крестообразный рисунок - опознавательный знак двойной спирали. Стало известно также, что нуклеотиды расположены друг от друга на расстоянии 0,34 нм, а на один виток спирали их приходится 10. Диаметр молекулы ДНК составляет около 2 нм. Из рентгеноструктурных данных, однако, было не ясно, каким образом цепи удерживаются вместе в молекулах ДНК.
Картина полностью прояснилась в 1953 г., когда американский биохимик Дж. Уотсон и английский физик Ф. Крик , исследуя структуру молекулы ДНК, пришли к выводу, что сахарофосфатный остов находится на периферии молекулы ДНК, а пуриновые и пиримидиновые основания - в середине. Предложена в 1953 Дж. Уотсоном и Ф. Криком.-«Чтобы придти к этому правильному решению, потребовалось найти такую конфигурацию, которая была бы стереохимически наиболее выгодной и в то же время не противоречила бы данным рентгено-структурного анализа» (Дж. Уотсон). Создание модели было подготовлено работами М. Уилкинса и Р.Франклин (получивших в 1950—52 высококачеств. рентгенограммы ДНК), Л. Полинга (создавшего в 1951 теорию, позволявшую предсказывать вид рентгенограмм для разл. спиральных структур), А. Тодда и его сотрудников (выяснивших в 1952 природу химич. связей между нуклеотидами, из к-рых построена ДНК), Э. Чаргаффа (установившего в 1947—50 соотношение азотистых оснований в ДНК). У. — К. м. позволила предсказать возможный механизм полуконсервативной матричной редупликации ДНК, общий принцип кодирования и транскрипции генетич. информации, нек-рые мол. механизмы мутационного процесса. Позднее в многочисл. исследованиях осн. положения и следствия из У.— К. м. получили эксперим. подтверждение. Уточнения коснулись более точного описания геометрич. параметров и конформационных возможностей двойной спирали при разл. условиях. В связи с обнаружением значит, конформационной подвижности структуры ДНК не раз поднимался вопрос о степени соответствия У.— К. м. структуре нативной ДНК. Предлагались др. гипотетич. модели ДНК, напр. неперевитая зигзагообразная модель ДНК, имеющая на границах поворотов-зигзагов право- и левозакрученные участки полинуклеотидных цепей. Существование левозакрученной, т. е. Z-ДНК, на отд. участках генома подтверждено экспериментально в работах А. Рича. Тем не менее нет оснований сомневаться в том, что У.— К. м. в осн. чертах правильно описывает структуру ДНК не только in vitro, но и in vivo. Создание У.— К. м. послужило мощным толчком к развитию мол. биологии, начало к-рой нередко датируют 1953.
д) Кодовая система ДНК.
Генетический код — это система записи информации о последовательности расположения аминокислот в белках с помощью последовательного расположения нуклеотидов в и-РНК.
Св-ва ген. кода:
1) Код триплетен. Это означает, что каждая из 20 аминокислот зашифрована последовательностью 3 нуклеотидов, называется триплетом или кодоном.
2) Код вырожден. Это означает, что каждая аминокислота шифруется более чем одним кодоном (исключение метиотин и триптофан)
3) Код однозначен — каждый кодон шифрует только 1 аминоксилоту
4) Между генами имеются «знаки препинания» (УАА,УАГ,УГА) каждый из которых означает прекращение синтеза и стоит в конце каждого гена.
5) Внутри гена нет знаков препинания.
6) Код универсален. Генетический код един для всех живых на земле существ.
Вопрос № 28.
ЗАКОНОМЕРНОСТИ НАСЛЕДОВАНИЯ.
а) Менделирующие признаки человека.
Хорошо известно явление так наз. множественного аллелизма, при к-ром ряд генов последовательно доминирует один над другим. Признаки, наследование к-рых подчиняется перечисленным закономерностям, принято называть менделирующими (по имени Г. Менделя).
У человека менделирующими признаками являются, напр., альбинизм (отсутствие пигментации, вызываемое рецессивным геном; встречается у всех человеческих рас с частотой 1 на 20— 30 тыс. новорожденных), цвет глаз, характер волос (курчавые или гладкие), групповые отличия по различным факторам в крови и др. Законам Менделя подчиняются и гены, обусловливающие наследственные болезни человека.
б) Независимое комбинирование неаллельных генов. 3-й закон Менделя (определение).
Свободное комбинирование при оплодотворении всех типов половых клеток приводит к реализации в потомстве всех возможных комбинаций генов. Возможность независимого комбинирования неаллельных генов определяется тем, что они находятся в разных парах хромосом. Одновременно и независимо протекающие по всем парам хромосом явления расщепления обеспечивают все возможные комбинации неаллельных генов между собой. Однако вскоре стало очевидно, что число неаллельных генов, свойственных любому виду растений и животных, должно превышать число присущих им пар хромосом. Поэтому неаллельные гены, находящиеся в одной и той же паре хромосом, должны наследоваться совместно
(3-й закон Менделя смотри выше)
в) Сцепленное наследование признаков (определение).
Сцепленное наследование — феномен скоррелированного наследования определённых состояний генов, расположенных в одной хромосоме.
Явление совместного наследования генов, локализованных в одной хромосоме, называется сцепленным наследованием, а локализация генов в одной хромосоме — сцеплением генов. Сцепленное наследование генов, локализованных в одной хромосоме, установил Морган.
г) Наследование признаков, сцепленных с полом (определение).
Наследование, сцепленное с полом — наследование какого-либо гена, находящегося в половых хромосомах. Наследование признаков, проявляющихся только у особей одного пола, но не определяемых генами, находящимися в половых хромосомах, называется наследованием, ограниченным полом.
Наследованием, сцепленным с X-хромосомой, называют наследование генов в случае, когда мужской пол гетерогаметен и характеризуется наличием Y-хромосомы (XY), а особи женского пола гомогаметны и имеют две X-хромосомы (XX). Таким типом наследования обладают все млекопитающие (в том числе человек), большинство насекомых и пресмыкающихся.
Наследованием, сцепленным с Z-хромосомой, называют наследование генов в случае, когда женский пол гетерогаметен и характеризуется наличием Z-хромосомы (ZW), а особи мужского пола гомогаметны и имеют две Z-хромосомы (ZZ). Таким типом наследования обладают все представители класса птиц.
Если аллель сцепленного с полом гена, находящегося в X-хромосоме или Z-хромосоме, является рецессивным, то признак, определяемый этим геном, проявляется у всех особей гетерогаметного пола, которые получили этот аллель вместе с половой хромосомой, и у гомозиготных по этому аллелю особей гомогаметного пола. Это объясняется тем, что вторая половая хромосома (Y или W) у гетерогаметного пола не несет аллелей большинства или всех генов, находящихся в парной хромосоме.
Таким признаком гораздо чаще будут обладать особи гетерогаметного пола. Поэтому заболеваниями, которые вызываются рецессивными аллелями сцепленных с полом генов, гораздо чаще болеют мужчины, а женщины часто являются носителями таких аллелей.
д) Полигенное наследование как механизм наследования количественных признаков.
Термин 'полигенного наследования' используется для обозначения наследования количественных признаков, черт, которые зависят от множества генов, а не одна. в дополнение к с участием нескольких генов, полигенных наследования и смотрит на роль окружающей среды в развитие кто-то.
Один легко понять например полигенных наследования по высоте. люди не просто короткий или высокий, они имеют различные высоты, которые идут вдоль спектра. кроме того, высота зависит также от окружающей среды; кто-то родился с высокими генов может стать короткий из-за недоедания или заболевания, например, в то время как кто-то родился с короткими генов может стать высокий путем генной терапии. основные генетики, очевидно, не будет достаточно, чтобы объяснить широкое разнообразие человеческих высот, но полигенных наследования показано, как несколько генов в сочетании с окружающей средой человека может влиять на фенотип кто-то, или внешности.
Цвет кожи является еще одним примером полигенных наследство, как и многие врожденные заболевания, потому что полигенных наследования настолько сложна, она может быть очень поглощающих и разочарование специальности. исследователи могут бороться идентифицировать все гены, которые играют роль в частности фенотип, а также определить места, где таких генов может пойти не так. однако, как только исследователям сделать больше узнать об обстоятельствах, которые приводят к выражению определенных признаков, это может быть очень полезным опытом.