

Основи генетики і селекції
Основні закономірності спадковості
Генетика — наука про закономірності спадковості і мінливості. Спадковістю називають властивість організмів повторювати в ряді поколінь подібні ознаки і забезпечувати специфічний характер індивідуального розвитку в певних умовах середовища. Завдяки спадковості батьки і потомки мають подібний тип біосинтезу, який визначає подібність хімічного складу тканин, характеру обміну речовин, фізіологічних функцій, морфологічних ознак та інших особливостей. Внаслідок цього кожний вид організмів відтворює себе із покоління в покоління.
Мінливість — це явище, певною мірою протилежне спадковості, і виявляється в тому, що у будь-якому поколінні
окремі особини чимось відрізняються і одна від одної, і від своїх батьків. Відбувається це тому, що властивості й ознаки кожного організму — це результат взаємодії двох причин: спадкової інформації і конкретних умов зовнішнього середовища, які можуть впливати як на зміну спадкових задатків, так і на варіабельність виявлення їх.
Генетика як наука виникла для задоволення практичних потреб. Під час розведення домашніх тварин і культурних рослин людина здавна використовувала гібридизацію, тобто схрещування організмів, які належали до різних видів, порід, сортів і відрізнялися один від одного якими-небудь ознаками. Порівнюючи гібриди з вихідними формами, практики давно помітили деякі особливості успадкування ознак. Ч. Дарвін надавав великого значення вивченню закономірностей спадковості та мінливості і встановив, що вони лежать в основі еволюції органічного світу.
На вивченні генетичних закономірностей грунтується селекція, тобто створення нових і поліпшення існуючих порід домашніх тварин, сортів культурних рослин, а також мікроорганізмів, які використовують у фармацев-тичній промисловості, медицині і народному господарстві.
Велике значення має генетика для медицини і ветеринарії, оскільки багато хвороб людини і тварин спадкові і для лікування їх або запобігання потрібні генетичні дослідження.
Основні закономірності успадкування властивостей і ознак були відкриті Г. Менделем (1822—1884). Однак його дослідження не відразу були належно оцінені і залишалися мало відомими до 1900 р., коли водночас три дослідники (Г. де Фріз у Голландії, К. Корренс у Німеччині і Е. Чермак в Австрії) незалежно один від одного вдруге відкрили закони спадковості, сформульовані Г, Менделем. Цю дату вважають датою створення експериментальної генетики.
Під час вивчення закономірностей успадковування зазвичай схрещують організми, які відрізняються один від одного альтернативними, тобто контрастними виявами ознаки. Наприклад, можна взяти насіння гороху (саме його брав Г. Мендель) жовтого і зеленого кольорів (ознака — колір насіння), зморшкувате і гладеньке (ознака — форма насіння), із пурпуровим і білим забарвленням квіток (ознака — колір квітки), з високим і низьким стеблом (ознака — розмір стебла).
Кожна ознака організму визначається одним або кілько-магенами. Кожний ген може існувати в кількох формах (станах), які називають алелями (алеломорфними парами). Алелі гена розміщені в гомологічних хромосомах в одних і тих самих місцях (локусах) (мал. 19).
Гомозиготи і гетерозиготи. Якщо в обох гомологічних хромосомах містяться однакові алелі (наприклад, обидва кодують жовте забарвлення насіння або обидва — зморшкувату форму насіння), такий організм називають гомозиготним. Якщо ж алелі різні (наприклад, в одній із гомологічних хромосом алель кодує жовтий пігмент, а в іншій, гомологічній їй хромосомі — алель зеленого пігменту або один алель кодує гладеньку форму насіння, а другий — зморшкувату, то такий організм називають гетерозиготним. Іншими словами, зиготу, яка утворилася злиттям гамет з однаковими алелями певного гена, називають гомозиготою. Гетерозигота утворюється злиттям гамет, які несуть у собі різні алелі даного гена. Один і той самий організм може бути гомозиготним за одним (або кількома) генами (вв, ББ) і гетерозиготним за Іншим (іншими; Аа, Гг; див. мал. 19).
Генотип і фенотип. Сукупність спадкових факторів організму (генів, хромосом, мітохондрій, пластид) називають генотипом. Сукупність усіх ознак і властивостей організму, які є результатом взаємодії генотипу із зовнішнім середовищем, називають фенотипом. Ось чому організми з однаковим генотипом можуть відрізнятися один від одного залежно від умов розвитку та існування. Межі, в яких змінюються фенотипові вияви генотипу, називають нормою реакції.
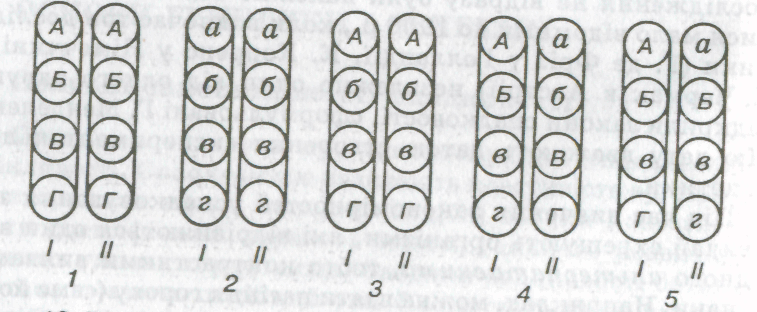
Мал. 19. Розміщення алелів гена (позначено однаковими літерами: великою — домінантний, малою — рецесивний) у гомологічних хромосомах:
1,2 — гомозигота відповідно за всіма домінантними і рецесивними алелями; 3 — гомозигота за частиною домінантних (АА, ГГ) і рецесивних (бб. вв) алелів; 4 — гетерозигота за всіма парами алелів; 5 — гомозигота за ББ- і вв- ознаками і гетерозигота за Аа- I г-ознакамм; /, // — групи зчеплення
Гібридологічний метод. Основні закономірності успад-ання були відкриті Г. Менделем. Він досяг успіхів у
своїх дослідженнях, застосувавши новий, розроблений ним метод, який дістав назву гібридологічного аналізу. Суть гібридологічного методу вивчення спадковості полягає в тому, що про генотип організму судять за ознаками (фенотипом) потомків, отриманих внаслідок певних схрещувань.
Метод ґрунтується на перелічених нижче положеннях
1. Враховують не весь різноманітний комплекс ознак батьків і гібридів, а аналізують спадковість за окремими ознаками та їхніми виявами.
2. Проводять точний кількісний облік спадкування кожного стану ознаки не лише в першому поколінні від схрещування, а й у наступних поколіннях. Гібридологічний метод широко застосовують у науці і практиці.
Об'єктом дослідження Г. Мендель вибрав горох, який має багато сортів, що відрізняються альтернативними виявами ознак. Вибір об'єкта виявився вдалим, оскільки спадкування ознак у гороху відбувається досить чітко. Гороху зазвичай властиве самозапилення, хоча можливе й перехресне запилення. У самозапильної рослини все потомство є потомством одного організму, тобто становить так звану чисту лінію, а здатність до перехресного запилення полегшує гібридизацію. Це дало Г. Менделю змогу проаналізувати потомство як кожної окремої особини, так і в результаті гібридизації. Перш ніж приступити до експериментів, Г. Мендель кілька років перевіряв чистоту сорту (гомозиготність), а впевнившись у цьому, розпочав експеримент.
Г. Мендель проаналізував закономірності спадковості як у разі, коли батьківські організми відрізнялися за альтернативним виявом однієї ознаки (моногібридне схрещування), так і в разі, коли вони відрізнялися за альтернативними виявами кількох ознак (ди-, три, полігібридне схрещування). Відповідно до рівня розвитку науки того часу Г. Мендель не міг пов'язати спадкові фактори з певними структурами клітини. Нині встановлено, що гени містяться в хромосомах, тому для пояснення закономірностей Менделя ми виходитимемо із сучасних цитологічних Уявлень про матеріальних носіїв спадковості.
Моногібридне схрещування. Перший закон Менделя. У дослідах Г. Менделя у разі схрещування сортів гороху з жовтим і зеленим насінням все потомство (тобто гібриди першого покоління) виявилися з жовтим насінням. ричому не мало значення, з якого саме насіння (жовтогочи зеленого) виросли материнські (батьківські) рослини. Отже, обоє батьків однаковою мірою здатні передавати свої ознаки потомству.
Аналогічні результати було отримано і в дослідах, в яких до уваги бралися інші ознаки. Так, у разі схрещування сортів із гладеньким і зморшкуватим насінням все потомство мало гладеньке насіння. У разі схрещування рослин з пурпуровими і білими квітками у всіх гібридів були лише пурпурові пелюстки квіток і т. д.
Виявлена закономірність дістала назву першого закону Менделя, або закону одноманітності гібридів першого покоління. Стан (алель) ознаки, який виявляється в першому поколінні, дістав назву домінантного; стан (алель), який у першому поколінні гібридів не виявляється, було названо рецесивним.
"Задатки" ознак (за сучасною термінологією — гени) Г. Мендель запропонував позначати літерами латинської абетки. Алелі, які належать до однієї пари станів ознаки, позначають однією й тією самою літерою, але домінантний алель — великою, а рецесивний — малою. Алель пурпурового забарвлення квіток слід позначати, наприклад, А, алель білого кольору квіток — а, алель жовтого кольору насіння — В, алель зеленого кольору насіння — 6 і т. д. Згадаймо, що кожна клітина тіла має диплоїдний набір хромосом. Усі хромосоми парні, а алелі гена містяться в гомологічних хромосомах. Отже, в зиготі завжди є два алелі і Генотипові формулу за будь-якою ознакою слід записувати двома літерами.
Особину, гомозиготну за домінантним алелем, слід записувати АА, за рецесивним — аа, гетерозиготну — Аа. Експериментально доведено, що рецесивний алель виявляє себе лише у гомозиготному стані, а домінантний — як у гомозиготному (АА), так і в гетерозиготному (Аа).
Гени розміщені у хромосомах. Отже, внаслідок мейозу гомологічні хромосоми (а з ними алелі гена) розходяться в різні гамети. Однак оскільки у гомозиготи обидва алелі однакові, всі гамети несуть один і той самий алель, тобто гомозиготна особина дає лише один тип гамет.
Досліди
щодо схрещування запропоновано записувати
у вигляді схем. Домовлено батьків
позначати буквою Р
особин
першого покоління — F1
(від
лат. filiale
— діти), особин другого покоління — Рг
І
т. д. Схрещування позначають знаком
множення (х), генотипову формулу
материнської особини
![]() записують першою, а батьківської
записують першою, а батьківської
![]() — другою. У першому рядку
записують
генотипові формули батьків, у другому
— типи їхніх гамет, у третьому — генотипи
першого покоління
і.т.д.
— другою. У першому рядку
записують
генотипові формули батьків, у другому
— типи їхніх гамет, у третьому — генотипи
першого покоління
і.т.д.
Для прикладу запишемо схему моногібридного схрещуання гомозиготного гороху з пурпуровими і білими квітками:
Р АА х аа
Гамети А а
F1 Аа
Оскільки у першого батька лише один тип гамет (А), а в другого також один тип гамет (а), можливе лише одне сполучення — Аа. Всі гібриди першого покоління виявляються одноманітними: гетерозиготними за генотипом і домінантними за фенотипом.
Отже, перший закон Менделя, або закон одноманітності гібридів першого покоління, у загальному вигляді можна сформулювати так: у разі схрещування гомозиготних особин, які відрізняються одна від одної за однією ознакою, яка має альтернативні (контрастуючи) прояви, усе потомство у першому поколінні одноманітне як за фенотипом, так і за генотипом.
Другий закон Менделя. У разі схрещування гетерозиготних гібридів першого покоління між собою (самозапилення або споріднене схрещування) у другому поколінні з'являються особини як з домінантними, так і з рецесивними станами ознак, тобто виникає розщеплення, яке відбувається в певних відношеннях. Так, у дослідах Г, Менделя на 929 рослин другого покоління виявилося 705 з пурпуровими квітками і 224 з білими. У досліді, в якому враховувався колір насіння, із 8023 насінин гороху, отриманих у другому поколінні, 6022 було жовтих і 2001 зелених, а із 7324 насінин, для яких враховувалася форма насінини, було отримано 5474 гладеньких і 1850 зморшкуватих.
Узагальнивши фактичний матеріал, Г. Мендель дійшов висновку, що у другому поколінні 75 % особин мають домінантний стан ознаки, а 25 % — рецесивний (розщеплення 3 : 1). Ця закономірність дістала назву другого закону Менделя, або закону розщеплення.
Згідно з цим законом та з використанням сучасної термінології можна дійти таких висновків: а) алелі гена, перебуваючи у гетерозиготному стані, не змінюють структуру один одного; б) під час дозрівання гамет у гібридів Утворюється приблизно однакове число гамет з домінантними і рецесивними алелями; в) у разі запліднення чоловічі і жіночі гамети, що несуть домінантні і рецесивні алелі, комбінуються вільно.
У разі схрещування двох гетерозигот (Аа), у кожної з яких утворюється два типи гамет (половина з домінантним алелем — А, половина — з рецесивним — а), слід очікувати чотири можливі поєднання. Яйцеклітина з алелем А може бути запліднена з однаковою часткою ймовірності як сперматозооном з алелем А, так і сперматозоо-ном з алелем а; а яйцеклітина з алелем а — сперматозооном з алелем А або з алелем а. Отримуються зиготи АА, Аа, Аа, аа, або АА, 2Аа, аа.
За зовнішнім виглядом (фенотипом) особини АА і Аа не відрізняються, тому дістають розщеплення у співвідношенні 3:1.
За генотипом особини розподіляються у співвідношенні 1АА : 2Аа : 1аа. Зрозуміло, що якщо від кожної групи особин другого покоління отримувати потомство лише шляхом самозапилення, то перша (АА) і остання (аа) групи (вони гомозиготні) даватимуть лише одноманітне потомство (без розщеплення), а гетерозиготні (Аа) форми даватимуть розщеплення у співвідношенні 3:1.
Отже, другий закон Менделя, або закон розщеплення, формулюється так: у разі схрещування двох гібридів першого покоління, які аналізують за однією ознакою з альтернативними проявами станів, у потомстві спостерігається розщеплення за фенотипом у співвідношенні З : 1 і за генотипом у співвідношенні 1:2: 1.
Статистичний характер законів генетики. Для встановлення деяких закономірностей біолог завжди має справу не з окремими поодинокими фактами або об'єктами дослідження, а з сукупністю фактів чи об'єктів. Кожний окремий об'єкт цієї сукупності характеризується своїми властивостями, оскільки він зазнає різноманітних впливів зовнішнього середовища.
Цих впливів може бути безліч, і за своєю дією вони настільки різноманітні, що виявити їх для кожного окремого випадку просто неможливо.
Незважаючи на це, всі разом взяті об'єкти виявляють певні статистичні закономірності (встановлені під час вивчення великого числа об'єктів), і біолог може передбачати наслідки масового явища загалом. Стосовно ж окремого факту або об'єкта сукупності можна говорити лише про ймовірність того, що він матиме місце, характеризуватиметься тими чи іншими властивостями.
Усі явища в природі можна поділити на випадкові і якономірні. У разі закономірних явищ за явищем А відбуватиметься явище В, у разі випадкових — у відповідь на явище А може відбутися не тільки явище В, а й С, D> тощо.
Саме тому в потомстві гібридів фактична кількість, яку отримують у досліді, не завжди відповідає очікуваній, оскільки генетичні співвідношення свідчать лише про ймовірність того, що в разі моно гібридного схрещування у другому поколінні 3/4 особин мають бути з домінантним станом ознак і 1/4 — з рецесивним. У разі малої кількості потомків фактичні числа можуть дуже відхилятися від очікуваних. Проте, як випливає з теорії ймовірності, чим більший фактичний матеріал, тим він більше наближається до очікуваного відношення. Приклад розв'язування задач із цього розділу див. у додатку — задача 4.
Гіпотеза "чистоти гамет". Поява серед гібридів другого покоління особин з рецесивним станом ознак дала Г. Менделю змогу зробити висновок про те, що "задатки", які визначають рецесивний стан ознаки, в гетерозиготному організмі не зникають, а лише пригнічуються. Оскільки передавання станів ознаки до потомків здійснюється через гамети, то було сформульовано гіпотезу чистоти гамет. Згодом ця гіпотеза отримала цитологічне обґрунтування. Ми знаємо, що у соматичних клітинах диплоїдний набір хромосом. В однакових місцях (локусах) гомологічних хромосом містяться алелі гена. Якщо це гетерозиготна особина, то в одній із гомологічних хромосом є домінантний алель, у другій — рецесивний. Під час утворення статевих клітин відбувається мейоз, і в кожну з гамет потрапляє галоїдний набір хромосом. У ньому, як відомо, всі хромосоми непарні, і природно, в гаметі може бути або домінантний, або рецесивний алель. Гамети залишаються "чистими" лише з якимось одним алелям, носієм одного з альтернативних станів ознаки.
Аналізуючи схрещування. Про генотип організму, який виявляє рецесивний стан ознаки, можна судити за його фенотипом. Справді, якби цей організм був гетерозиготним, то у нього виявився б домінантний стан ознаки. Отже, якщо виявляється рецесивний стан ознаки, то організм обов'язково має бути гомозиготним за рецесивним алелем. Розглянемо це на прикладі. З досвіду відомо, що у кролів чорний колір шерсті (ген А) домінує над білим (ген a). Якщо кріль має білий колір шерсті, то природно, що хоча б один із алелів гена, який кодує колір шерсті, має бути а. Другий же алель не може бути А, бо в такому разі
кріль (згідно з першим законом Менделя) був би чорним. Отже, і другий алель у цій парі — а, тобто генотип білого кроля стосовно кольору — аа.
Інша річ у особин, які виявляють домінантні стани ознаки. Гомозиготна і гетерозиготна особини в цьому разі за фенотипом не відрізняються. Наприклад, якщо кріль має чорне забарвлення шерсті, за фенотипом неможливо визначити його генотип. Він може бути як гомозиготним, так і гетерозиготним. Для визначення генотипу проводять аналізуючи схрещування. Суть цього схрещування полягає в тому, що особину, генотип якої потрібно з'ясувати, схрещують з особиною, гомозиготною за рецесивним станом ознак, після чого аналізують гібриди F
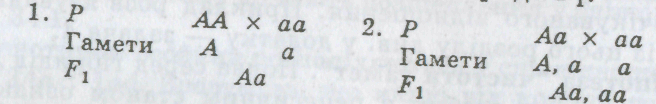
Як видно зі схеми, при аналізуючому схрещуванні в потомстві гомозиготної домінантної особини розщеплення немає, гетерозиготна ж особина дає розщеплення 1:1. Отже, за допомогою аналізуючого схрещування отримано відповідь на запитання про генотип особин, які виявляють у фенотипі домінантну ознаку.
Визначення генотипу має велике значення для проведення селекційних робіт у тваринництві і рослинництві (див. с. 113—118).
Проміжний характер успадкування. У своїх дослідах Г. Мендель мав справу з прикладами повного домінування, тому гетерозиготні особини в його дослідах вевідрізнялися від домінантних гомозигот. Однак у природі поряд з повним домінуванням часто трапляється неповне домінування (проміжне успадкування), тобто гетерозиготи мають власний фенотип. Так, у нічної красуні відомі два сорти: з червоними і білими квітками. Гібриди, що утворюються від схрещування цих сортів, мають рожеві квітки, тобто вони мають проміжний характер, У другому поколінні розщеплення за фенотипом відповідає розщепленню за генотипом, тобто 1 червона: 2 рожеві: 1 біла.
В андалузьких курей буває чорне і біле пір'я, а їхні гібриди мають "голубе" забарвлення. У разі схрещування великої рогатої худоби червоної і білої масті отримують гібриди чалої масті, тобто частина шерсті у них біла, а частина — червона і розташована вона на тілі тварини більш-менш рівномірно. Явище проміжного успадкування свідчить про те, що розвиток ознаки зумовлений взаємодією ферментів, які кодуються різними генами. Приклад розв'язування задач із цього розділу див. у додатку —
задача 5.
Відхилення від очікуваного розщеплення. Часом розщеплення у другому поколінні може відрізнятися від очікуваного. Як зазначалося раніше, причиною відхилення може бути мале число гібридів. Однак іноді однією з причин може бути наявність у генотипі деяких особин таких генів, які викликають їх загибель. Так, у разі схрещування жовтих мишей з чорними в потомстві з'являються жовті і чорні особини у співвідношенні 1 : 1. У разі схрещування між собою чорних мишей все потомство тільки чорне, в разі схрещування жовтих — у першому поколінні відбувається розщеплення у співвідношенні 2 жовтих: 1 чорна. Таке успадкування жовтого кольору пояснюють тим, що воно зумовлене домінантним алелям гена, але, мабуть, жовтий колір реалізується лише у гетерозиготному стані, про що свідчить аналіз усіх типів схрещування. Причина полягає в тому, що гомозиготні домінантні особини гинуть ще в ембріональному стані. Позначимо алель жовтого кольору А, а чорного — а, тоді

Подібний тип успадкування характерний для сірих каракульських овець, у яких внаслідок схрещування між собою спостерігається розщеплення у співвідношенні 2:1. Виявилося, що ягнята, гомозиготні за домінантним алелем сірого кольору, гинуть внаслідок недостатнього розвитку травної системи. Тому у господарствах, в яких розводять каракульських овець, незважаючи на те, що сірий смушок ціниться більше, сірих овець не схрещують з сірими плідниками, а схрещують сірих з чорними. Тоді втрат молодняка немає, розщеплення отримують у співвідношенні 1 :1.
Дигібридне і полігібридне схрещування. Організми різняться між собою за багатьма ознаками. Схрещування, за якого враховують відмінності за кількома ознаками, називають полігібрид ним. Найпростішим прикладом полі-гібридного схрещування є дигібрине, за якого у батьківських організмів береться до уваги відмінність за двома ознаками. Наприклад, два сорти гороху з різними кольорами і формою насіння. Перше покоління гібридів у цьому разі виявиться одноманітним, виявляться лише обидва домінантні алелі, незалежно від того, як ці ознаки були розподілені у батьків. Наприклад, якщо в одного з батьків було жовте і гладеньке насіння (ААВВ), а у другого — зелене і зморшкувате (ааЬЬ), то всі потомки за фенотипом виявляться подібними до першого з батьків. Однак якщо поєднання ознак у батьків було іншим, наприклад зелене і гладеньке насіння (ааВВ) в одного з батьків і жовте зморшкувате {ААЬЬ) — в другого, то у гібридів з'явиться одна ознака від одного, друга — від другого із батьків. Фактично ж в обох випадках одна й та сама закономірність — реалізація закону одноманітності у гетерозиготних форм.
Схематично ці схрещування можна записати так:
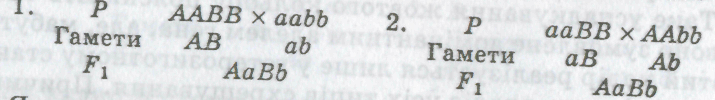
Як видно з наведених схем, у разі дигібридного схрещування дигомозиготна особина може бути: гомозиготною за домінантними (ААВВ) або рецесивними (ааЬЬ) алелями, за одним домінантним і одним рецесивним алелем (ааВВ, ААЬЬ). Особини можуть бути за одним із генів гомозиготними, а за другим — гетерозиготними (ААВЬ, ааВЬ, АаВВ, АаЬЬ), а також гетерозиготними за обома генами (АаВЬ) (див. мал. 19).
У другому поколінні було виявлено: жовтого гладенького насіння — 315, жовтого зморшкуватого — 101, зеленого гладенького — 108, зеленого зморшкуватого — 32.
Третій закон Менделя, або закон незалежного успадкування ознак. Вивчаючи розщеплення в разі дигібридного схрещування, Г. Мендель звернув увагу на таку обставину. Під час схрещування рослин з жовтим гладеньким (ААВВ) і зеленим зморшкуватим (ааЬЬ) насінням у другому поколінні з'являлися нові комбінації ознак: жовте зморшкувате (А_ЬЬ) і зелене гладеньке (ааВ_), яких не було у вихідних форм. Із цього спостереження Г. Мендель зробив висновок, що розщеплення за кожною ознакою (за кожною парою алелів) відбувається незалежно від другої ознаки (других пар алелів). У цьому прикладі форма насіння успадковувалась незалежно від його забарвлення. Ця закономірність дістала назву третього закону Менделя, або закону незалежного розподілу генів.
Третій закон Менделя формулюється так: у разі схрещування гомозиготних особин, які відрізняються за двома(або більше) ознаками, у другому поколінні (F2) спостерігаються незалежне успадкування І комбінування станів ознак, якщо гени, які їх визначають, містяться у різних парах хромосом. Це можливо тому, що під час мейозу розподіл (комбінування) хромосом у статевих клітинах під час їх дозрівання йде незалежно і може зумовити появу нащадків з комбінацією ознак, відмінних від батьківських і прабатьківських особин.
Оскільки кожна пара алелів розподіляється у гібридів незалежно від іншої пари, то в цьому прикладі у дигетерозиготної особини (АаВЬ) під час формування гамет алель А може виявитися в одній гаметі як з алелем Б, так і з алелем Ь. З такою самою ймовірністю і алель а може потрапити в одну гамету з алелем В або з алелем Ь. Отже, у дигетерозиготної особини утворюється чотири типи можливих комбінацій генів у гаметах: АВ, АЬ, аВ, аЬ. Всіх типів гамет буде порівну (по 25 %).
Це легко пояснити поведінкою хромосом під час мейозу. Для спрощення візьмемо гіпотетичний організм, який має всього дві пари хромосом. Назвемо їх першою і другою парами. Якщо цей організм гетерозиготний за обома генами, то одна із хромосом першої пари нестиме в собі алель А, друга — алель а; у другій парі хромосом одна із них нестиме алель В, друга — алель Ь. Після мейозу кожна гамета має по одній хромосомі з кожної пари. Негомо-логічні хромосоми під час мейозу можуть комбінуватися в будь-яких поєднаннях, тому хромосома з алелем А рівно-ймовірно може потрапити у гамету як з хромосомою з алелем В, так і з хромосомою з алелем Ь. Подібно може розподілитися і хромосома з алелем а: або з алелем В, або з алелем Ь.
Для запису схрещувань нерідко використовують спеціальні решітки, запропоновані англійським генетиком Р. Пеннетом (решітка Пеннета). Ними зручно користуватися під час аналізу полігібридних схрещувань. Принцип побудови решітки полягає в тому, що зверху по горизонталі записують гамети батьківської особини, зліва по вертикалі — гамети материнської особини, в місцях перетину — ймовірні генотипи потомства (мал. 20, 21).
Як видно з мал. 20, із 16 можливих комбінацій у другому поколінні в 9 реалізуються домінантні стани обох ознак (А_В__), у 3 — перша ознака має домінантний стан,
Друга — рецесивний (А__ЬЬ), ще в 3 — перша ознака має
Рецесивний, друга — домінантний стан (ааВ__) і, нарешті,
в одному — обидві ознаки мають рецесивний стан (ааЬЬ).
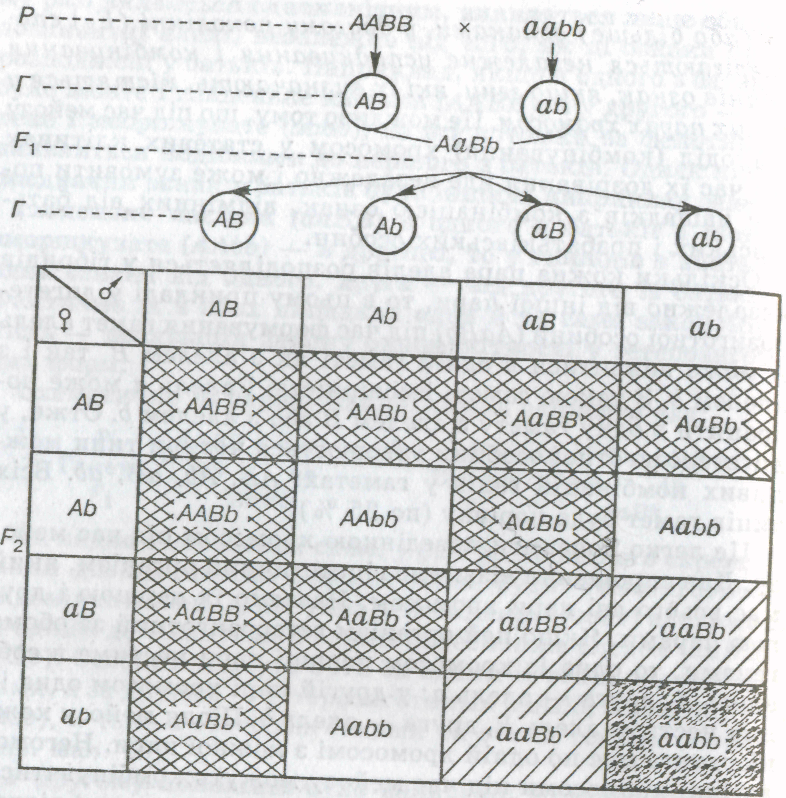
Мал. 20. Схема дигібридвого схрещування:
Р — батьки; Г — гамети; F1i F2г — гібриди відповідно першого і другого поколінь
ідбулося розщеплення за фенотипом у співвідношенні 9:3:3:1.
Якщо простежити розщеплення лише за забарвленням насіння, отримаємо: жовтих — 12/16, зелених — 4/16; якщо ж аналізувати лише за формою насіння, то виявиться гладеньких — 12/16, зморшкуватих — 4/16, тобто відоме розщеплення у співвідношенні 3:1. Це підтверджує незалежність успадкування ознак.
Якщо під час схрещування аналізують більш як дві ознаки, то кількість очікуваних комбінацій збільшиться. У разі три гібридного схрещування гетерозиготи утворюють по 8 типів гамет (див. мал. 21), які дають 64 комбінації. Якщо всі можливі комбінації записати у вигляді решітки Пеннета, а потім проаналізувати її, то виявиться,

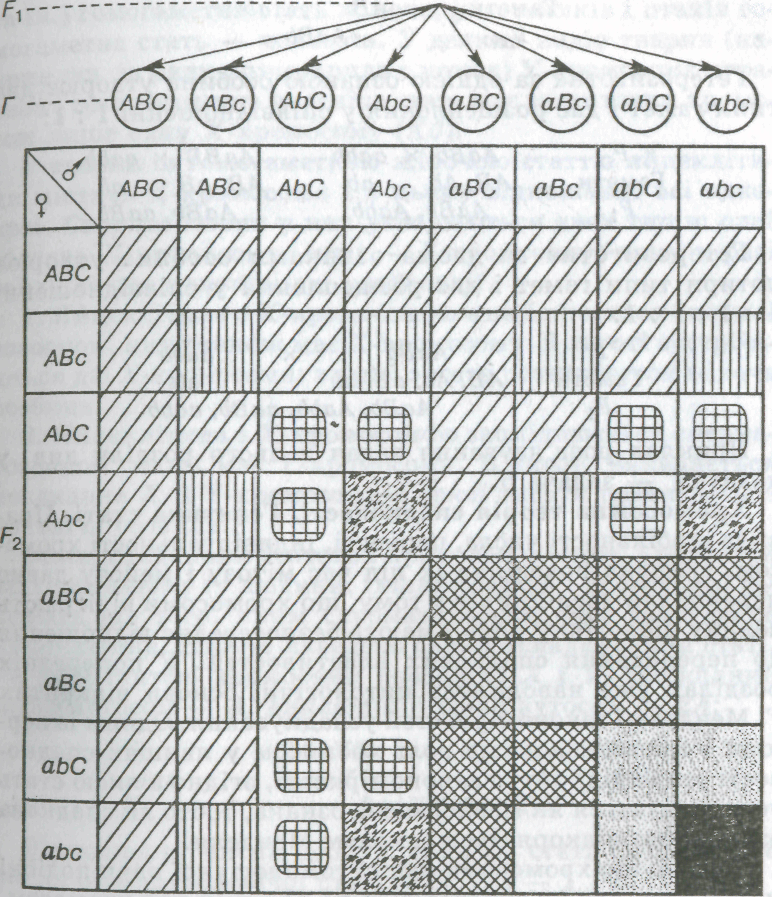
Мал. 21. Схема тригібридного схрещування (позначення ті самі, Що й на мал. 20)
Що розщеплення за фенотипом відбулося у співвідношенні 27 : 9 : 9 : 9 : 3 : 3 : 3 : 1.
Розщеплення за фенотипом у загальній формі можна виразити формулою (3 + 1)", де п ~ кількість ознак, які взято для аналізу під час схрещування.
у разі аналізуючого схрещування число типів потомків вказує на число типів гамет, що утворює особина, генотип
якої аналізують. Під час схрещування домінантної гомозиготної особини розщеплення не спостерігається, бо вона утворює один тип гамет з домінантними алелями:
Р ААВВ х ааЬЬ
Гамети АВ аЬ
F1 АаВЬ
Гетерозиготна за однією ознакою особина утворює два типи гамет і дає розщеплення у співвідношенні 1:1:
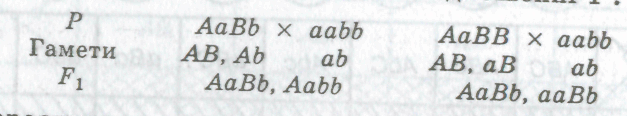
Гетерозиготна за двома ознаками особина утворює чотири типи гамет і дає розщеплення у співвідношенні 1:1:1:1:
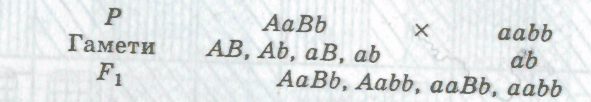
Приклад розв'язування задач з цього розділу див. у додатку — задача 6.
Хромосомна теорія спадковості. Генетика статі. Правила стабільності числа, парності, індивідуальності хромосом, складна поведінка їх під час мітозу і мейозу давно переконали дослідників у тому, що хромосоми відіграють велику біологічну роль і мають безпосереднє відношення до передавання спадкових властивостей. У попередніх розділах уже наводились цитологічні основи відкритих Г. Менделем закономірностей успадкування. Одним із перших і вагомих доказів ролі хромосом у явищах спадковості стало відкриття закономірності, згідно з якою стать успадковується як менделююча ознака, тобто як спадкова ознака, що підкоряється законам Менделя.
Відомо, що хромосоми однієї гомологічної пари подібні між собою, але це справедливо не для всіх пар хромосом. Порівнянням хромосомних наборів із соматичних клітин жіночої і чоловічої особин в одній парі хромосом виявлено відмінності, хоча в однієї із статей і ці хромосоми однакові, їх називають X (ікс)-хромосомами. У другої статі — одна така сама Х-хромосома, а друга відрізняється за своєю будовою. Вона названа У (ігрек)-хромосомою. Цю пару прийнято називати статевими хромосомами, а всі пари хромосом, ідентичні у чоловічої і жіночої особин, — ауто-сомами. Статеві (X і У) хромосоми відрізняються не лише за морфологією, а й за інформацією, яка міститься в них.
Сполучення статевих хромосом у зиготі визначає стать майбутнього організму. Стать з однаковими статевими хромосомами (XX) називають гомогаметною, з різними (ХУ) — гетерогаметною.
у багатьох видів тварин (а також у людини, дрозофіли та ін.) гомогаметна стать жіноча, у метеликів і птахів гомогаметна стать — чоловіча. У деяких видів тварин (наприклад, із ряду прямокрилих комах) У-хромосома втрачена і гетерогаметна чоловіча стать має із статевих хромосом лише одну Х-хромосому (Х0).
У тварин із гомогаметною жіночою статтю яйцеклітини містять Х-хромосоми і в цьому відношенні всі однакові. Сперматозоони у них утворюються ддвох типів: одні містять Х-хромосому, інші — У-хромосому, тому в разі запліднення можливі дві комбінації.
1. Яйцеклітина з Х-хромосомою запліднюється сперма-тозооном, який також має Х-хромосому. В зиготі зустрічаються дві Х-хромосоми; з такої зиготи розвивається жіноча особина.
2. Яйцеклітина з Х-хромосомою запліднюється сперма-тозооном, який має У-хромосому. В зиготі виявляється поєднання X- і У-хромосом; із такої зиготи розвивається чоловіча особина.
Отже, поєднання статевих хромосом у зиготі і розвиток статі організму (у людини, інших ссавців і дрозофіли) залежать від того, яким сперматозооном буде запліднена яйцеклітина. Нижче наведено схему успадкування статі, на якій статеві хромосоми позначені X і У, галоїдний набір ауто сом — А, диплоїдний набір аутосом — 2А:
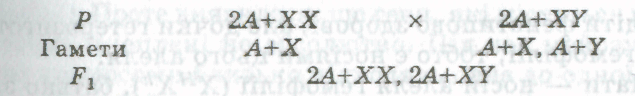
Відношення числа особин жіночої статі (2А+ХХ) до числа особин чоловічої статі (2А+ХУ) — 1:1.
Сперматозоонів з X- і з У-хромосомою приблизно однакові кількості, тому особин обох статей народжується приблизно порівну.
Ознаки, які успадковуються через статеві X- і У-хромосоми, називають зчепленими зі статтю. У людини ознаки, які успадковуються через У-хромосому, характерні лише для особин чоловічої статі, а успадковані через Х-хромосому — як для однієї, так і для другої статі. Особини жіночої статі можуть бути як гамо-, так і гетерозиготними за генами, які локалізуються в Х-хромосомі, а рецесивні алелі
у них виявляються лише в гомозиготному стані. Оскільки в особин чоловічої статі лише одна Х-хромосома, то всі локалізовані в ній гени, навіть рецесивні, відразу ж вияв-ляються у фенотипі.
Коли записують схеми передавання зчеплених зі статтю ознак, то у генетичних формулах поряд із символами генів записують і статеві хромосоми.
У людини зчепленими зі статтю успадковуються гемофілія (порушене згортання крові), дальтонізм та ін. Ген, який контролює нормальне згортання крові (Н), і його алельна пара — "ген гемофілії" (h) — містяться в Х-хро-мосомі. Якщо жінка гетерозиготна за цією ознакою (ХНХН), то гемофілія у неї не виявляється. У чоловіка лише одна Х-хромосома, отже, його генотип може бути ХНУ і ХЛУ. Оскільки У-хромосома не несе ген, який визначає механізм згортання крові, то стан його здоров'я за цією ознакою визначатиме Х-хромосома: ХИУ — здоровий, ХНУ — хворий на гемофілію.
Гетерогаметні організми, які в статевих хромосомах диплоїдного набору мають лише один алель гена, називають гемізиготними.
Успадкування гемофілії можна проілюструвати прикладами.
1. Мати має кров з нормальним процесом згортання і гомозиготна за цією ознакою (ХИХН), батько хворіє на гемофілію (ХНУ):
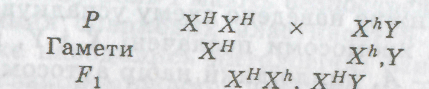
Всі діти фенотипові здорові, але дочки гетерозиготні за
геном гемофілії, тобто є носіями цього алеля.
2. Мати — носій алеля гемофілії (ХИХН), батько здоровий (ХпУ):
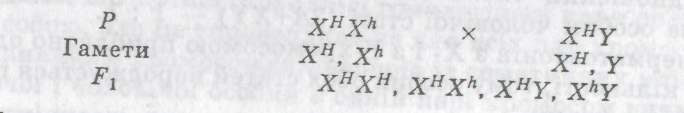
У цьому разі всі дочки за фенотипом здорові, але половина з них є носіями, серед синів — половина хворих на гемофілію.
Природно, що рецесивний алель гемофілії у гетерозиготному стані може зберігатись у жінок навіть упродовж кількох поколінь, доки знову не виявиться у когось із осіб чоловічої статі. Дочка, хвора на гемофілію, може народиться у родині, де мати гетерозиготна за цією ознакою (носій), а батько хворий на гемофілію. Оскільки ця хвороба рідкісна, таке поєднання малоймовірне.
Аналогічно успадковується дальтонізм, тобто аномалії зору, коли людина не розрізняє кольорів. Нормальне сприйняття кольорів зумовлене домінантним алелем, який локалізується в Х-хромосомі, його рецесивний алель у гомозиготному стані призводить до виникнення дальтонізму. Звідси зрозуміло, чому дальтонізм значно частіше трапляється У чоловіків: у них лише одна Х-хромосома (гемі-зигота), і якщо в ній міститься рецесивний алель дальтонізму, він обов'язково виявиться. У жінок дві Х-хромосоми, вони можуть бути як гетерозиготними за цим геном, так і гомозиготними, але тільки в останньому разі вони страждатимуть на дальтонізм. Приклад розв'язування задач з цього розділу див. у додатку — задача 7.
Зчеплене успадкування. Вище було наведено приклади успадковування ознак через статеві хромосоми. Носіями спадковості є також і аутосоми. Оскільки генів значно більше, ніж хромосом, то природно, що кожна хромосома несе велику кількість генів, які успадковуються разом. Гени, розміщені в одній хромосомі, називають групою зчеплення (див. мал. 19). У кожного виду організмів число груп зчеплення дорівнює числу пар хромосом, тобто у дрозофіли їх 4, у гороху — 7, у кукурудзи — 10 і т. д.
Встановлений Г. Менделем третій закон незалежного успадкування і комбінування ознак справедливий лише тоді, коли гени, які визначають ці ознаки, містяться в різних парах гомологічних хромосом (належать до різних груп зчеплення). Проте виявилося, що гени, які містяться в одній хромосомі, зчеплені не абсолютно. Під час мейозу гомологічні хромосоми близько підходять одна до одної, переплітаються, відбувається перехрещування і вони можуть обмінюватися своїми ділянками (мал. 22). Обмін ділянками може відбутися в будь-якому місці. Чим далі один від одного розміщені гени в одній хромосомі, тим частіше між ними може відбутися перехрещування і обмін ділянками.
У дрозофіли алелі генів, що визначають довжину крил (нормальні А і короткі а) і забарвлення тіла (сіре В і чорне Ь), розміщені в одній парі гомологічних хромосом, тобто належать до однієї групи зчеплення.
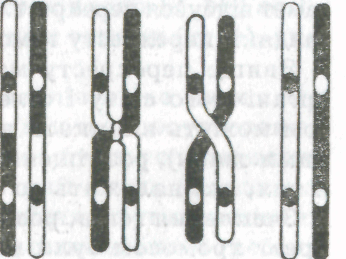 22.
Схема перехрещування хромосом
22.
Схема перехрещування хромосом
Якщо схрестити гомозиготних мух за рецесивними (ab//ab) і домі-
Нантними( AB//AB) проявами ознаки, то в першому поколінні
все потомство буде гетерозиготним (AB//AB). Для з ясуванпя,
які саме гамети утворює особина першого покоління, потрібно провести аналізуюче схрещування — схрестити гібридну особину першого покоління з гомозиготною за рецесивними проявами особиною, тобто чорною короткокрилою.
Якщо два гени різних алельних пар розміщені у різних хромосомах, то у дигетерозиготи (як уже зазначалося) чергується чотири типи гамет: 25 % гамет АВ, 25 % АЬ, 25 % аВ, 25 % аЬ, тобто при аналізуючому схрещуванні має бути чотири типи потомків: сірі довгокрилі, сірі короткокрилі, чорні короткокрилі, чорні довгокрилі у співвідношенні 1:1:1:1.
Однак взяті для прикладу гени містяться в одній групі зчеплення. Обидва домінантні алелі розміщені в одній із гомологічних хромосом, а рецесивні — у другій гомологічній хромосомі, тому гени А і В не можуть комбінуватися незалежно один від одного. У разі абсолютного зчеплення обох генів слід очікувати лише на два типи гамет — 50 % АВ і 50 % аЬ і появу у другому поколінні лише двох типів потомків: сірих довгокрилих і чорних короткокрилих. Так і трапляється в разі схрещування гібридного самця з гомозиготною рецесивною самкою. Пояснюють це тим, що у самця дрозофіли перехресту хромосом не спостерігається. Проте якщо схрестити гібридну самку з гомозиготним рецесивним самцем, то з'являться всі чотири типи мух, але з новим співвідношенням ознак, тобто сірих короткокрилих (ааВ_) і чорних довгокрилих (АЬЬ) буде значно менше, ніж сірих довгокрилих (А_В_) і чорних короткокрилих {ааЬЬ). Нові поєднання ознак виникають лише там, де під час утворення гамет відбувся перехрест. Отже, зчеплення генів порушується завдяки перехресту гомологічних хромосом.
Явище перехресту має велике значення для еволюції органічного світу і селекції. Завдяки перехресту хромосом можуть виникати нові комбінації генів (і кодованих ними ознак), розміщених не тільки у різних хромосомах, а й тих, які належать до однієї групи зчеплення.
Зчеплення генів, розміщених в одній хромосомі, і перехрест хромосом були відкриті генетиком Т. Морганом.
Нині вивчено групи зчеплення для багатьох видів організмів: дрозофіли, миші, кукурудзи, томатів. Відомі групи зчеплення для всіх хромосом людини.
Основні положення хромосомної теорії спадковості. Відкриті Т. Морганом закономірності, підтверджені і поглиблені згодом на численних об'єктах, відомі під загальною назвою хромосомна теорія спадковості. Нижче наведено основні її положення.
1. Гени містяться у хромосомах. Кожна пара хромосом є групою зчеплення генів. Число груп зчеплення у кожного виду дорівнює числу пар хромосом.
2. Кожний ген у хромосомі займає певне місце (локус). Гени в хромосомі розміщені лінійно.
3. Між гомологічними хромосомами можуть відбуватися перехрест і обмін генами.
4. Частота перехресту прямо пропорційна відстані між генами. Чим далі розміщені гени, тим частіше відбувається перехрест. Приклад розв'язування задач з цього розділу див. у додатку — задача 8.
Новоутворення під час схрещування. Розвиток будь-яких ознак організму — це наслідок складних взаємодій між генами, точніше, між продуктами їхньої діяльності — білками, ферментами. Часто для виявлення ознаки потрібна взаємодія ферментів, які кодуються двома різними генами. Так, у разі схрещування двох сортів горошку запашного з білими квітками гібриди першого покоління виявилися не білими, а червоно-фіолетовими. Новоутворення зумовлене тим, що для синтезу пігменту потрібні два різних ферменти, які кодуються різними домінантними алелями. У разі схрещування таких гібридних рослин відбувається розщеплення у співвідношенні 9 : 7. Генетичним аналізом доведено, що генотипи вихідних сортів горошку з білими квітками були ААЬЬ і ааВВ. Гібриди першого покоління були гетерозиготними (АаВЬ). У другому поколінні всі рослини з домінантними алелями обох генів мали червоно-фіолетове забарвлення, а рослини, що мали лише якийсь один із домінантних алелів (А_ЬЬ і ааВ_) і були гомозиготними за рецесивними генами (ааЬЬ), виявилися однотипними, білими.
Часом кожний із домінантних алелів може мати самостійний вияв, проте разом вони дають новоутворення. Класичним прикладом цього може бути успадкування форми гребеня у курей. Домінантний алель А визначає розвиток трояндоподібної форми гребеня, другий домінантний алель В зумовлює розвиток горохоподібного гребеня,
рецесивні алелі (аЬ) кодують листкоподібний гребінь, a наявність двох домінантних алелів (АВ) забезпечує новоутворення — горіхоподібний гребінь.
Типи взаємодії генів. Взаємодію генів і алелів можна подати такою схемою:
Домінування виявляється в тих випадках, коли один алель гена повністю приховує наявність іншого (за фенотипом при повному домінуванні гібриди подібні лише до одного з батьків). Проте, очевидно, найчастіше наявність рецесивного алеля якимось чином виявляється, і зазвичай доводиться зустрічатися з неповним домінуванням різного ступеня.
 Це
пояснюється тим, що домінантний алель
відповідає за активну форму білкафермента,
а рецесивні алелі часто детермінують
ті самі білки-ферменти, але зі зниженою
ферментативною активністю. Це явище і
реалізується у гетерозиготних форм
у вигляді неповного домінування.
Це
пояснюється тим, що домінантний алель
відповідає за активну форму білкафермента,
а рецесивні алелі часто детермінують
ті самі білки-ферменти, але зі зниженою
ферментативною активністю. Це явище і
реалізується у гетерозиготних форм
у вигляді неповного домінування.
Наддомінування полягає в тому, що у домінантного алеля в гетерозиготному стані іноді спостерігається сильніший прояв, ніж у гомозиготному стані.
Кодомінування — прояв у гетерозиготному стані ознак, що детермінуються обома алелями. Наприклад, кожен з алелів кодує певний білок, І в гетерозиготному організмі синтезуються вони обидва. У такому разі біохімічними дослідженнями можна встановити гетерозиготність без проведення аналізуючого схрещування. Цей метод застосовують у медико-генетичних консультаціях для виявлення гетерозиготних носіїв генів, що зумовлюють хвороби обміну. За типом кодомінування у людини успадковується четверта група крові.
Комплементарними називають взаемодоповнювальні гени, коли для формування ознаки потрібна наявність кількох неалельних (як правило, домінантних) генів. Цей тип успадкування дуже поширений у природі, У запашного горошку колір віночка квітки зумовлений наявністю двох домінантних генів (А_В_), при відсутності одного з них квітки білі. Тому при схрещуванні рослин з генотипами АаЬЬ і ааВВ, що мають білі віночки, у першому поколінні рослини мають забарвлення, у другому поколінні розщеплення відбувається у співвідношенні 9 забарвлених (А_В_) до 7 н-езабарвлених (ЗА_ЬЬ, ЗааВ_ і ХааЬЬ).
Подібно успадковується колір у папуг-нерозлучників (А_ЬЬ — жовті, ааВ_ — блакитні, А_В_ — зелені, ааЬЬ — білі).
Молекула гемоглобіну крові дорослої людини містить чотири поліпептидних ланцюги, сполучені із залізовмісним гемом. Для синтезу молекули гемоглобіну необхідна наявність двох комплементарних генів.
Епістаз — протилежна комплементарній взаємодія генів. Під епістазом розуміють пригнічення неалельним геном (епістатичним) дію іншого гена, який називають гіпостатичніш. Наприклад, у курей домінантний алель гена С зумовлює розвиток пігменту, але домінантний алель іншого гена / є його супресором, "пригнічувачем". Тому кури, навіть ті, що мають домінантний алель гена кольору, наявності супресора виявляються білими. Особини, що мають у генотипі гени І_С_, — білі, а з генотипом ііС__ — пігментовані. Отже, білий колір курей може бути зумовлений як відсутністю домінантного алеля гена пігментації, так і наявністю домінантного алеля гена, що зумовлює явище епістазу (пригнічення) гена пігментації.
Полімерія. Різні домінантні неалельні гени можуть впливати на одну й ту саму ознаку, підсилюючи її прояв. Такі гени називають однозначними, або полімерними, а ознаки, що ними визначаються, — полігенними. У цьому разі два або більше домінантних алелів однаковою мірою впливають на розвиток однієї й тієї самої ознаки. Полімерні гени прийнято позначати однією літерою з цифровими індексами (Аи А2,А3).
Генотип як цілісна система. Наведені приклади новоутворень у разі схрещування свідчать про те, що розвиток організму зумовлений складною взаємодією генів. Можливо, що розвиток будь-якої ознаки пов'язаний з дією багатьох генів. Крім того, виявлено залежність кількох ознак від одного гена. Наприклад, у вівса забарвлення лусочок і довжина остюка насіння визначаються одним геном. У дрозофіли ген білого кольору очей одночасно впливає на колір тіла і внутрішніх органів, довжину крил, зниження плодючості, зменшення тривалості життя. Не виключено, що кожний ген є одночасно геном основної дії для "своєї" ознаки і модифікатором для інших ознак. Отже, фенотип — це результат взаємодії генів усього генотипу із зовнішнім середовищем в онтогенезі особини. Склалася ця цілісна система у процесі еволюції органічного світу, виживали лише ті організми, в яких взаємодія генів дала найсприятливішу реакцію в онтогенезі.
Цитоплазматична спадковість. Основна роль у передаванні спадкових властивостей належить хромосомам. З ними пов'язані закономірності, відкриті Г. Менделем і Т. Морганом. Проте багато органел цитоплазми містить ДНК (пластиди, мітохондрії). Вони здатні до авторепродукції і з їхньою ДНК може бути пов'язана передача по-захромосомної, цитоплазматичної спадковості.
Є сорти ротиків, нічної красуні та деяких інших рослин, у яких поряд із зеленими листками трапляються строкаті з білими плямами — ділянками, позбавленими хлорофілу. Ознака строкатості передається тільки по материнській лінії, через пластиди.
У кукурудзи є сорти з чоловічою стерильністю, яка передається винятково через цитоплазму жіночих статевих клітин. Сорти кукурудзи, що характеризуються чоловічою стерильністю, широко використовують у сільському господарстві під час лінійних схрещувань. Внаслідок відсутності власного пилку у цих ліній неможливе самозапилення, тому завдяки перехресному запиленню отримують гібрид з підвищеною врожайністю (див. "Гетерозис"). Якою саме органелою передається чоловіча цитоплазматична стерильність у кукурудзи, ще не встановлено.
Значення генетики для медицини. Закономірності успадкування ознак єдині для всіх організмів. Вивчення їх у людини пов'язано з певними труднощами. Цілком зрозуміло, що на людині неможливо ставити експерименти. Повільна зміна поколінь, невелика кількість дітей у кожній родині також гальмують вивчення генетики людини.
Генетику людини вивчають такими методами: генеалогічним, близнюковим, цитологічним.
Генеалогічний метод полягає у складанні родоводу. Цим методом встановлено, наприклад, що карий колір очей домінує над блакитним, наявність ластовиння — над його відсутністю, здатність краще володіти правою рукою над здатністю краще володіти лівою рукою тощо. Цим методом встановлено також здатність передавати у спадок деякі хвороби.
Близнюковий метод полягає у вивченні розвитку ознак
близнят. Відомо, що у людини близнята бувають двох категорій: різнояйдеві та однояйцеві. Різнояйцеві розвиваються з різних яйцеклітин, кожна з яких запліднена "своїм" сперматозооном. Такі близнята можуть бути як однієї, так і різних статей. Вони схожі між собою не більше, ніж звичайні брати і сестри. Оскільки вони розвиваються в однакових умовах, то всі відмінності між ними зумовлені генотипом. Однояйцеві близнята розвиваються з однієї зиготи, яка розділилася на два фрагменти на певних етапах розвитку зародка. Вони обов'язково однієї статі і настільки подібні один до одного, що їх важко розпізнають навіть батьки. Всі відмінності між ними зумовлені факторами середовища, а подібність — генотипом. Цей метод часто дає змогу встановити, яка роль спадковості і середовища у розвитку різних ознак, а також захворювань. Наприклад, на кір хворіють як однояйцеві, так і різнояйцеві близнята, отже, ця хвороба цілком зумовлена факторами середовища, тобто залежить від потрапляння в організм збудника хвороби. На туберкульоз і дифтерію можна захворіти в разі потрапляння в організм збудника, але в ризику захворіти велику роль відіграє і генотип. Зазвичай, якщо на таку хворобу захворів один із однояйцевих близнят, то часто захворює і другий. У різнояйцевих такої за-кономірностд немає. Групи крові цілком зумовлені спадковістю, вони завжди однакові в однояйцевих близнят.
Цитологічний метод полягає у вивченні хромосомних комплексів. З цією метою зазвичай використовують лейкоцити у мазках крові. Цим методом встановлено низку мутацій, які призводять до тяжких захворювань. Наприклад, коли в диплоїдному наборі хромосом виявиться одна зайва хромосома із 21-ї пари (47 замість 46), то це призведе до розвитку хвороби Дауна. Такі хворі мають малий розмір голови, вузький розріз очей, плоске обличчя і різко знижений інтелект. Виявлено і багато інших хвороб, по-в язаних з порушенням кількості і структури хромосом.
Встановлено, що близько 4 % немовлят мають спадкові хвороби або вади в будові тіла. В разі ранньої діагностики спадкової хвороби можна домогтися усунення або послаблення її впливу на розвиток організму. Деякі хвороби, пов язані із зміною числа хромосом, лікують гормонами. озвитку деяких спадкових хвороб можна запобігти спеціальною дієтою. Приблизно в одному випадку на десять | місячновонароджених дитина має спадкову хворобу, повязану з відсутністю ферментів, які необхідні для засвоєння молочного цукру. Якщо діагноз буде поставлено несвоєчасно і дитину годувати грудним молоком, у неї розвинеться недоумкуватість. Якщо ж діагноз поставлено вчасно і дитина переведена на штучне вигодовування, вона розвиватиметься нормально. З цього прикладу можна зробити висновок, що поява спадкової хвороби не фатальна. На жаль, слід зазначити, що при цьому хоча хвороба й не фатальна, однак це призводить до зростання кількості небажаних алелів у популяції. Звідси випливає важливість завдання медицини й охорони здоров'я — вивчення спадкових хвороб, розроблення методів їх профілактики, діагностики і лікування. Особливо великого значення слід надавати розробці методів профілактики спадкових хвороб.
Нині у великих містах нашої країни створено медико-генетичні консультації. Це одна із форм профілактики спадкових хвороб, куди можуть звернутися за порадою здорові батьки дітей, що мають якісь дефекти, або хворі на спадкові хвороби; батьки, один з яких хворіє на спадкову хворобу; здорові батьки, що мають родичів з дефектом розвитку або хворих на спадкові хвороби; подружжя, які перебувають у кровноспорідненому шлюбі, та ін.
Профілактика спадкових хвороб спрямована також на ослаблення дії мутагенних факторів: зменшення опромінення природними і штучними джерелами, зниження вмісту хімічних мутагенів у навколишньому середовищі (промислових відходів, речовин побутової хімії, пестицидів). Слід нагадати, що хімічні речовини (алкоголь, нікотин, наркотики), потрапляючи в організм, можуть змінювати нормальну послідовність нуклеотидів у ДНК і структурі хромосом, порушувати процес розподілу хромосом під час мейозу. Це може призвести до виникнення спадкових хвороб і аномалій розвитку.
Контрольні запитання і завдання
1. Схарактеризуйте генетику як науку.
2. Дайте визначення явищ спадковості і мінливості.
3. Який зв'язок генетики з сільським господарством і медициною?
4. Які гени називають алельними? Як виникли алельні гени?
5. Дайте визначення понять "гомозигота", "гетерозигота", "генотип", "фенотип".
6. У чому суть гібридологічного методу?
7. Які основні закономірності спадкування встановив Г. Мендель?
8. Які стани ознаки називають домінантними і рецесивними?
9. Дайте характеристику моногІбридного та дигібридяого схрещування.
10 Скільки типів гамет утворює гомозиготна особина у разі моногІбридного та дигіОридного схрещування?
11 Скільки типів гамет утворюють гетерозиготи за однією ознакою?
12 Скільки типів гамет утворює гетерозиготна особина за двома ознаками?
13. Скільки утвориться генотипів і фенотипів у другому поколінні в разі мово- та дигібридиого схрещування?
14. Поясніть, чому особини, які мають домінантні стани ознаки, можуть мати різний генотип, а рецесивні — одноманітні за генотипом.
15. Що таке аналізуюче схрещування?
16. Чому закономірності спадкування мають статистичний характер?
17. Дайте цитологічне пояснення розщеплення ознак у другому
ПОКОЛІННІ.
18. Сформулюйте закон незалежного спадкування ознак та дайте йому цитологічне пояснення.
19. Побудуйте решітку Пеннета для чотиригібридного схрещування.
20. Який хромосомний механізм генетичного визначення статі?
21. Як успадковуються гемофілія і дальтонізм?
22. Дайте цитологічне пояснення зчепленого успадкування.
23. Дайте характеристику цитоплазматичної спадковості.
24. У чому складність вивчення спадковості у людини? Дайте характеристику методів вивчення спадковості у людини.
Закономірності мінливості
Живий організм завжди перебуває в тісному взаємозв'язку з навколишнім середовищем, під впливом факторів якого виявляється одна з найважливіших властивостей організму — його мінливість. Саме вона забезпечує різноманітність організмів у процесі еволюційного розвитку. Якщо в процесі мінливості відбувається зміна генотипу, то ознака, що виникла у відповідь на п,ю зміну, успадковується і виявляється у наступних поколіннях. Таку мінливість називають генотиповою, мутаційною, спадковою, иевизначеною. Якщо ж зміни організму, що виникли У відповідь на вплив навколишнього середовища, не торкнулися генотипу, а лише викликали більш-менш значні зміни фенотипу, то йдеться про фенотипову, модифікаційну, неспадкову, визначену мінливість. Такі зміни не успадковуються. Порівняння цих двох типів мінливості наведено в табл. 2.
Вплив умов зовнішнього середовища на розвиток і виявлення ознак. Модифікаційна мінливість. Розглянемо
Таблиця 2. Порівняння модифікаційної та мутаційної мінливостей
Зміни, що виникли внаслідок модифікаційної мінливості |
Зміни, що виникли внаслідок мутаційної мінливості |
Потомками не успадковуються
Пов'язані тільки з фенотипом і на генотип не впливають З'являються масово
Формуються під прямим впливом середовища, мають спрямований характер
Мають пристосувальний характер |
Потомками успадковуються Пов'язані з перебудовою в спадковому апараті (генах, хромосомах) З'являються лише в окремих особин Мають неспрямований характер; одні й ті самі мутації можуть виникати в результаті дії різних факторів і, навпаки, під впливом одного фактора виникають різні мутації Пристосувального характеру не мають; можуть бути шкідливими, нейтральними, корисними |
приклад. У кімнатної рослини первоцвіту китайського за звичайних умов за температури від 15 до 20 °С квітки мають червоне забарвлення. Однак, якщо рослину з червоними квітками перенести у приміщення з підвищеною вологістю і температурою ЗО—35 °С, то нові квітки матимуть білий колір, а в разі повернення рослини у попередні умови знову з'являться червоні квітки. Це означає, що одні й ті самі спадкові властивості виявляються по-різному залежно від умов, в яких відбувається реалізація цих властивостей.
Кролі гімалайської породи мають білу шерсть і лише вуха, ніс, лапи і хвіст чорного кольору (іноді бурого). Якщо на спині кролика виголити або вищипати частину білої шерсті й утримувати його на холоді, нова шерсть виросте чорною. Навпаки, на частинах тіла, які мають темну шерсть, за підвищеної температури можуть відрости білі волосини (мал. 23). Подібний результат отримано в дослідах на сіамських котах. У них переважає жовтувате забарвлення, але вуха, ніс, лапи, хвіст чорного кольору. Змінюючи температуру середовища, вдається і у таких котів домогтися темного забарвлення шерсті на всіх частинах тіла або білої на тих його частинах, які зазвичай мають чорний колір. Було проведено серію експериментів над рослинами, що належали до 120 видів. Кожний екземпляр рослин розрізали на дві однакові частини. Одну частину вирощували в умовах теплого м'якоїго клімату, а другу — високо в горах. Результа-
Мал. 23. Феаотнпова зміна забарвлення шерсті гімалайського кролика під впливом температури:
/
— кролик, який виріс за звичайної
температури середовища (близько 20 °С);
2
—
кролик, який виріс за високої
температури середовища (близько 32 °С);
3 — кролик, у якого на спині виголена
частина шерсті і шкіра охолоджувалася
під стерильною пов'язкою; 4
—
кролик з пігментованою шерстю на
ділянці тіла, яка охолоджувалась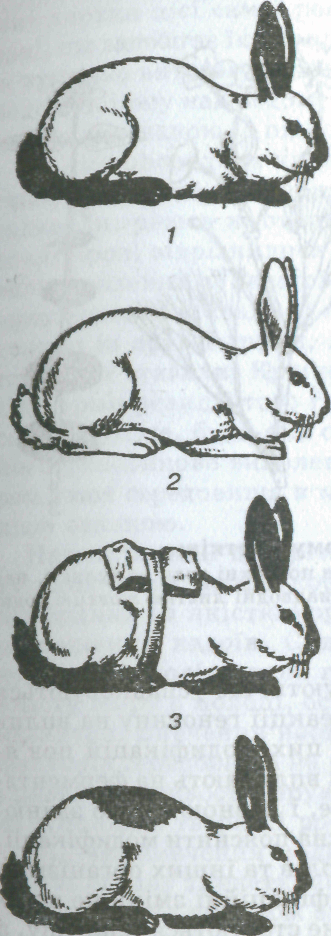
Мал. 24. Земляна груша:
1 — виросла на рівнині; 2 — виросла в горах (а — у тому самому масштабі, що й на
рівнині, б — у збільшеному масштабі)
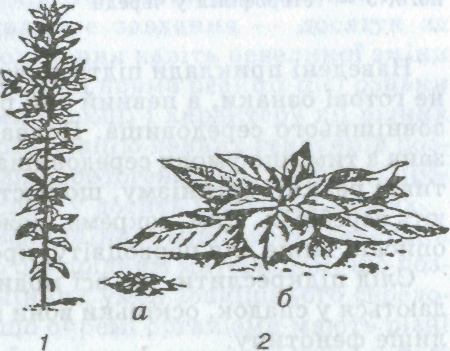
ти дослідів були такими (на прикладі земляної груші). Стебло земляної груші, яка виросла на рівнині, було високим, а вирощеної в горах — дуже низеньким і мало вигляд розетки, притисненої до землі (мал. 24). Така зміна форми стебла — результат пристосування до суворих умов високогір'я. Якщо насіння обох рослин висівати в однотипних умовах, вони дадуть рослини з однаковим фенотипом незалежно від того, яким був фенотип їхніх батьків. Отже, під впливом зовнішніх умов зміни у фенотипі не спричинюють змін генотипу. Навіть на одній рослині листки, які виросли у різних Умовах, мають різну форму (явище гетерофілії, мал. 25).
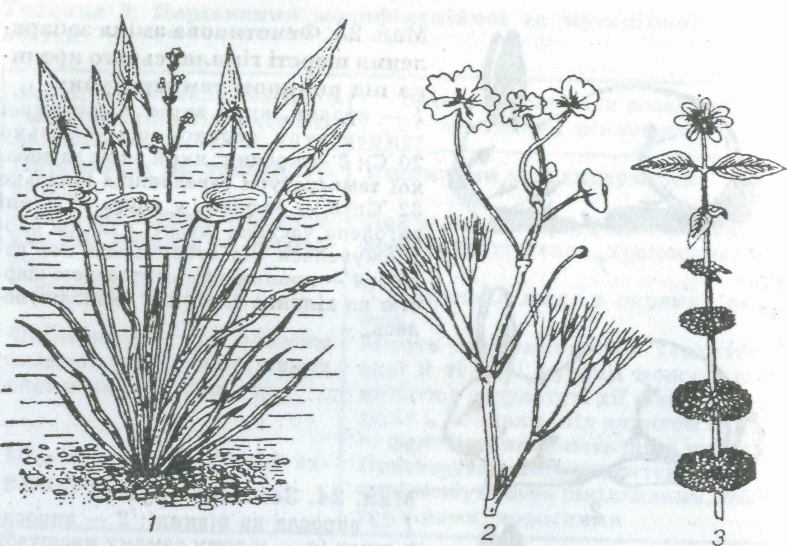
Мал. 25. Вплив середовища на форму листків:
1 — стрілолист (у воді листки лінійні, на поверхні води — кулясті, над подою — стрілоподібні); 2 — підводні і надводні листки жовтцю водяного; 3 — гетерофілія у череди
Наведені приклади підтверджують, що успадковуються не готові ознаки, а певний тип реакції генотипу на вилив зовнішнього середовища. Поява цих модифікацій пов'язана з тим, що умови середовища впливають на ферментативні реакції організму, що росте, і певною мірою змінюють перебіг їх. Цим, зокрема, можна пояснити модифікації, описані вище для первоцвіту, кролів та інших організмів.
Слід підкреслити, що всі модифікаційні зміни не передаються у спадок, оскільки вони не стосуються генотипу, а лише фенотипу.
Діапазон, в якому залежно від умов зовнішнього середовища може змінюватися фенотипове виявлення генотипу, називають нормою реакції. Норма реакції, яка лежить в основі модифікаційної мінливості, формувалась історично в результаті природного добору. Виживали ті організми, в яких під впливом середовища відбувалися фенотипові зміни, що сприяли їх існуванню. Отже, норма реакції відповідає умовам життя, є пристосувальною.
Дуже демонстративні у цьому плані листки жовтцю водяного. Ця рослина росте як у воді, так і на березі. Іноді лише частина рослини занурена у воду. Над водою листки мають широкі пластинки із слабко розрізаними края-
ми; листки цієї самої рослини під водою дуже розчлено-вані Щ° запобігає їхньому пошкодженню течією води. Якщо
ж зумисно витримувати частину листкової пластинки під водою, а іншу над водою, то надводна частина листка буде нерозчленованою, а підводна набуде глибоких вирізів.
На чистопородних кроленятах одного приплоду було поставлено такий експеримент. Частину кроленят щедро годували, решта ж отримували мінімальний раціон. Дорослі кролі відрізнялися між собою. Проте як би не годували кроленят із першої групи, більшими за певну величину вони не ставали, і навпаки, як би мало не годували тварин із другої групи, меншими за певні розміри вони також не ставали. Крайні величини росту і тут залежать від норми реакції того генотипу, який тварини дістали у спадок. Отже, будь-яка ознака зумовлена генотипом, але його фенотипове виявлення може змінюватися під впливом умов середовища в межах норми реакції генотипу за цією ознакою.
Норма реакції для різних ознак може бути широкою і вузькою. Так, якщо корову годувати певною кількістю підібраних за якістю кормів, то можна досягти значного підвищення надоїв. Складніше завдання — досягти за допомогою поліпшення годування навіть невеликої зміни вмісту жиру в молоці, оскільки норма реакції цієї ознаки незначна. Вивчення норми реакції і характеру дії різних факторів середовища на реалізацію певних генотипів розкриває можливості керування онтогенезом і, зокрема, продуктивністю тварин і рослин у досить широких межах.
Під час вивчення модифікаційної мінливості генетики мають справу з генетично однорідним матеріалом, що розвивається під впливом змінних умов зовнішнього середовища. Це спричинює те, що окремі організми мають різні параметри (кількісні) тієї чи іншої ознаки. Отже, у разі вивчення закономірностей модифікаційної мінливості завдання зводиться до вивчення закономірностей у масі випадкових явищ. Для цього використовують методи математичної статистики.
Статистичні закономірності модифікаційної мінливості. Варіаційний ряд. Зміни багатьох ознак піддаються кількісному вивченню. Так, зріст і маса тіла людини однієї статі і вікУ, довжина листків якогось дерева, маса насіння квасолі та інших рослин, молочність великої рогатої худоби, несучість курей і багато інших ознак можна оцінити кількісно.
Як приклад розглянемо число реберець на черепашці морського молюска серцевидки їстівної. Підрахунком
Мал.
26. Графік мінливості (варіаційна
крива) числа реберець на черепашці
молюска серцевидки їстівноїреберець
у 100 особин було виявлено, що їх число
коливається від 19 до 26. Якщо підрахувати,
скільки особин має те чи інше число
реберець на черепашці і записати цічисла
у порядку зростання (або спадання), то
дістанемоваріаційний ряд: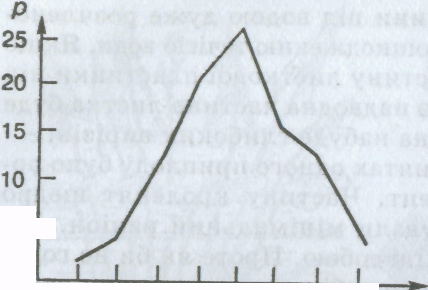
Число реберець
(розмір ознаки v) 19 20 21 22 23 24 25 26
Кількість особин з даним
числом реберець
(кількість варіант р) 2 4 11 21 26 16 13 7
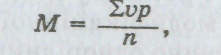
де
![]() —
знак суми; V
—
варіанти (розмір ознаки); р
—
частота зустрічальності; п
—
загальна кількість досліджених
особин у варіаційному ряді.
—
знак суми; V
—
варіанти (розмір ознаки); р
—
частота зустрічальності; п
—
загальна кількість досліджених
особин у варіаційному ряді.
Обчислимо за цією формулою середню величину для наведеного прикладу:
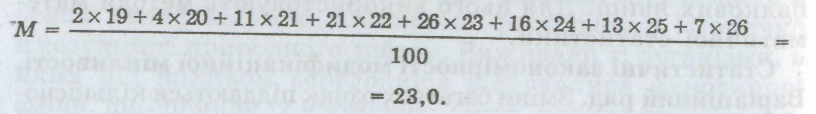
Отже, середнє число реберець на черепашці молюска дорівнює 23. У цьому разі воно збігається з найчастіше зустріваною варіантою (26 разів із 100). Такий збіг не обов'язковий, проте ці величини зазвичай мало відрізняються одна від одної.
Якими ж причинами зумовлений такий розподіл варіант у варіаційному ряді? Це наслідок неоднорідності факторів зовнішнього середовища, які викликають зміни в межах норми реакції. На організми діють як сприятливі, так і несприятливі фактори. Поєднання їх випадкове. В основі випадкових явищ лежать специфічні закономірності, які виявляються лише в разі великої кількості варіант. Ці закономірності називають статистичними, або закономірностями множинних причин. Вони вивчені математично з позицій теорії ймовірності. За цією теорією, поєднання тільки сприятливих або тільки несприятливих умов трапляється рідко. Тому більшість організмів мають величину ознаки середню або близьку до середньої. Організмів із значними відхиленнями ознак від середніх величин у природі мало. Це можна чітко простежити за варіаційною кривою.
Генотипову (або спадкову) мінливість прийнято поділяти на комбінативну і мутаційну.
Комбінативна мінливість пов'язана з отриманням нових комбінацій генів у генотипі. Досягається це в результаті трьох процесів: незалежного розходження хромосом під час мейозу; випадкового сполучення їх під час запліднення; обміну генами в гомологічних хромосомах у разі перехресту. Самі по собі гени внаслідок цього не змінюються, проте нові їхні сполучення зумовлюють появу організмів з новими фенотипами.
Ч. Дарвін встановив, що багато сортів культурних рослин і порід свійських тварин з'явилися завдяки гібридизації предків. Він надавав великого значення комбіна-тивній мінливості, оскільки їй, поряд з добором, належить важлива роль в утворенні нових форм як у природі, так і в народному господарстві.
Слід підкреслити, що біологічне значення статевого розмноження полягає у можливості отримання нових комбінацій спадкової інформації. Саме тому статеве розмноження так поширене в природі, й, отже, ця мінливість має велике значення для еволюції.
Мутаційна мінливість. Процес мейозу супроводжується не лише комбінуванням генотипу, а й значною реорганізацією окремих генів — мутаціями. Мутації виникають раптово, стрибкоподібно й іноді різко відрізняють організм від вихідної форми.
Рослинникам і тваринникам спадкові зміни були відомі Давно. Деякі з них описав Ч. Дарвін у праці "Про мінливість домашніх тварин і культурних рослин (1868). Мутаційній мінливості присвятили свої праці С. І- Коржинський (1899) і де Фріз (1901). Останньому належить термін "мутації".
Мутації відомі у всіх класів тварин, рослин, грибів, бактерій, вірусів. З мутаційною мінливістю пов'язана еволюція — процес утворення нових видів, сортів і порід. За характером змін генетичного апарату розрізняють мутації, зумовлені як зміною числа хромосом (геномні) та їхньої структури (хромосомні), так і змінами молекулярної структури гена (генні, або точкові, мутації).
Збільшення числа хромосом, кратне гаплоїдному набору (Зл, 4л і т. д.), називають поліплоїдією. Виникнення поліплоїдів пов'язане з порушенням мейозу. Поліплоїдію широко використовують у селекції. Детальніше вона буде розглянута нижче.
Зміни в будові хромосом виникають у результаті того, що під час поділу клітини окремі ділянки хромосом можуть відриватися і губитися або приєднуватися до іншої гомологічної чи не гомологічної хромосоми. Це також спричинює зміни спадкових властивостей.
Більшість мутацій, з якими пов'язані еволюція органічного світу і селекція, — генні мутації. Вони є наслідком змін ділянок молекули ДНК, перестановки нуклеотидів, випадання окремих із них або вмонтування нового нукле-отиду.
Мутації можуть виникати у будь-яких клітинах, тому розрізняють мутації соматичні і генеративні (у статевих клітинах). Біологічне значення цих мутацій нерівнозначне і пов'язане з характером участі їх у розмноженні організмів.
Під час поділу соматичної клітини після виникнення мутації нові властивості передаються її потомкам. Тому в разі вегетативного розмноження нова ознака, що виникла внаслідок мутації в соматичній клітині, може зберігатися у потомків. Якщо у рослини мутаційні зміни відбулися в клітині, з якої утворюється брунька, а потім пагін, то останній нестиме нові властивості. Так, на кущі чорної смородини може з'явитися гілка з білими ягодами. Із соматичної мутації на яблуні Антонівці звичайній І. В. МічурІн вивів новий сорт — Антонівку півторафунтову.
Під час статевого розмноження ознаки, які з'явилися внаслідок соматичних мутацій, потомкам не передаються і в еволюції жодної ролі не відіграють. Проте в індивідуальному розвитку вони можуть впливати на формування ознаки: чим раніше (наприклад, до формування бласту-
ли) виникне соматична мутація, тим більшою виявиться ділянка, клітини якої несуть цю мутацію. Такі особини називають мозаїками. Мозаїками, наприклад, є люди, в яких різний колір правого і лівого ока, або тварини певної масті, в яких на тілі з'явились плями іншого кольору.
Якщо мутації відбуваються у клітинах, з яких розвиваються гамети, або у статевих клітинах, то нова ознака виявиться у найближчому або у наступних поколіннях. Спостереження довели, що майже всі мутації шкідливі для організму. Це пояснюють тим, що в організмі функціонування всіх органів добре збалансоване між собою і з зовнішнім середовищем. Порушення існуючої рівноваги зазвичай призводить до зниження життєздатності або до загибелі організму. Мутації, які негативно впливають на життєдіяльність, називають семілетальними, або напівлеталь-ними. Мутації, не сумісні з життям, називають летальними. Проте незначна частина мутацій може виявитись корисною. Такі мутації створюють матеріал для еволюції, а також для селекції цінних порід свійських тварин і культурних рослин. Можливі види мутацій подано нижче.
1. За змінами спадкового Зміни числа хромосом {геномні)
апарату Перебудова хромосом (хромосомні)
2. За характером клітин, Соматичні
в яких відбулися зміни Генеративні
3. За впливом на Шкідливі (летальні, напівлегальні,
Життєздатність стерильні)
Корисні (матеріал для добору)
Нейтральні
4. За причинами, що зумо- Спонтанні (природні)
вили зміни Індуковані
Частота мутацій. Мутація кожного окремого гена — явище досить рідкісне. Вважається, що у людини в середньому кожний окремий ген мутує з частотою 0,00005 на покоління. Проте оскільки генів дуже багато (у людини, можливо, кількасот тисяч), то та чи інша мутація трапляється не так рідко. Встановлено, що у дрозофіли близько 5 % гамет мають мутовані гени.
Накопичення мутацій у популяціях. Генетичний вантаж. Отже, зрозуміло, що в кожному поколінні мутуе значна кількість генів. Еволюція організмів пов'язана із заміною одних генотипів іншими завдяки природному добору, в основі якого лежить мінливість (мутації) і спадковість. Мутації, змінюючи гени, дають матеріал для перерозподілу генетичної інформації, а також для добору. Більшість мутацій шкідливі. Деякі з мутацій за тих умов, у яких існує дана популяція організмів, виявляються більш-менш нейтральними. Незначна частина мутацій якоюсь мірою корисна для життя організму, для існування виду. Домінантні мутації (алелі) починають контролюватися добором уже в першому поколінні, і їхня частота в популяції швидко зростає. І тут же добором усуваються організми, що є носіями домінантних летальних генів. Домінантні алелі, які частково знижують життєдіяльність або плодючість, деякий час зберігатимуться у популяції, але поступово і вони повністю зникають (елімінують). Рецесивні мутації можуть перебувати у популяції в прихованому стані. Вони починають контролюватися добором тільки після того, як досягнуть у популяції певної частоти прояву, почнуть переходити в гомозиготний стан. Гомозиготний організм за цим геном матиме інший фенотип, тобто відбудеться вищеплення мутації. Елімінація шкідливих рецесивних алелів відбувається значно повільніше, ніж домінантних, а повна елімінація шляхом добору, можливо, і не досягається. Нові мутації, як правило, рецесивні, у гетерозиготному стані не виявляються. Накопичення мутантних алелів створює комбінативну мінливість, яка зумовлює генетичну різноманітність природних популяцій. Отже, виникає генетичний вантаж. Для виду генетичний вантаж корисний, він забезпечує його існування і удосконалення. У разі зміни умов вид завжди має мобілізаційний резерв, або резерв спадкової мінливості.
Залежність мутаційного процесу від умов середовища. Експериментальне отримання мутацій. Мутації поділяють на спонтанні та індуковані. Спонтанними називають мутації, що виникають під впливом невідомих природних факторів, індукованими — мутації, спричинені спеціально спрямованими діями, які інтенсифікують мутаційний процес.
Фактори, що здатні індукувати мутаційний процес, дістали назву мутагенних. До мутагенних факторів належать деякі хімічні сполуки та види випромінювання.
Експериментально доведено, що з хімічних сполук мутагенну дію виявляють формалін, етиленімін, іприт та багато інших. У лабораторних умовах радіаційний мутагенез отримують під дією рентгенівських променів. Часто використовують гамма-промені, джерелом яких зазвичай є радіоактивний кобальт. Мутагенні фактори широко використовують у селекційній практиці, оскільки вони дають широкий спектр мутацій — матеріал для штучного добору.
Є всі підстави вважати, що причиною спонтанних мутацій є ті самі мутагенні фактори, які використовують і в експерименті. Останнім часом доведено також мутагенну дію вірусів. Спонтанні мутації — матеріал для природного добору.
Забруднення природного середовища мутагенами та Його наслідки. Життя сучасного суспільства пов'язане з широким застосуванням у промисловості, сільському господарстві і побуті багатьох хімічних сполук. Усе більше використовується атомна енергія. У зв'язку з цим необхідний контроль, щоб не допустити накопичення у навколишньому середовищі мутагенних факторів, які можуть загрожувати спадковості людини та інших організмів. Науковим комітетом ООН з радіації встановлено, що для людини дозою рентгенівських і гамма-променів, яка подвоює кількість природних мутацій, є (129.,.387) • 10~4 Кл/кг(50— 150 Р). Звідси зрозуміло, наскільки небезпечним є безконтрольне підвищення радіаційного фону навколишнього середовища, небезпечне не лише використання, а й випробування ядерної зброї.
Людство є свідком тяжких мутагенних наслідків хімічної війни США проти В'єтнаму з 1961 до 1971 р. Хромосомні мутації відмічаються упродовж трьох десятиліть не лише у дітей в'єтнамців, а й у дітей американських ветеранів, які побували у зонах хімічних атак в Індокитаї.
Закон гомологічних рядів спадкової мінливості (закон Вавилова). Відомо, що мутагенез відбувається в різних напрямах. Проте ця різноманітність підпорядкована певній закономірності, яку виявив у 1920 р. М. І. Вавилов. Порівнянням ознак різних сортів культурних рослин і близьких до них диких видів було виявлено багато спільних спадкових змін. Це дало М. І. Вавилову змогу сформулювати закон гомологічних рядів у спадковій мінливості: генетично близькі види і роди (пов'язані між собою єдністю походження) характеризуються подібними рядами у спадковій мінливості з такою закономірністю, Що, знаючи ряд форм у межах одного виду, можна передбачити існування паралельних форм інших видів і родів.
Проілюструємо це на прикладі (мал. 27). У колоскових злаків — м'якої і твердої пшениць і ячменю — відомі форми з довгими і короткими остюками і без остюків, а з опуклостями на їх місці. У вівса, пшениці та ячменю ко-

Мал. 27. Гомологічні ряди мінлицості за остюками:
1 — 4 — м'яка пшениця; 5-8 - тверда пшениця; 9-12 – шестирядний ячмінь
Форми з довгими(1,5,9) і короткими(2,6,10)_ острюками, здуті(3,7,11) і без острюків(4,8,12)
лоски бувають забарвлені в три основних кольори: червоний, чорний і білий.
Ще М. І. Бавилов зазначав, що гомологічні ряди часто виходять за межі родів і навіть родин. Короткопалість відмічена у багатьох рядів ссавців. Вона трапляється у великої рогатої худоби, овець, собак, людини. Альбінізм спостерігається у всіх класів хребетних.
Коли був сформульований закон гомологічних рядів, озима форма твердої пшениці ще не була відома. Однак існування її передбачалося. Через кілька років таку форму було виявлено в Туркменії.
У злаків — пшениці, ячменю, вівса, кукурудзи —існують голі і плівчасті зерна. Перша форма не була відома, але існування її передбачалось. І вона була знайдена.
Закон гомологічних рядів дає змогу передбачити можливість появи мутацій, ще не відомих науці, які можна використовувати у селекції для створення нових цінних для народного господарства форм.
Контрольні запитання і завдання
1. Дайте визначення генотипової і фенотипової мінливості.
2. Дайте порівняльну характеристику модифікаційної та мутаційної мінливості,
3. Що таке норма реакції? Наведіть приклади.
4. Чи успадковуються ознаки, які виникли у межах норми реакції?
5. Дайте характеристику статистичних закономірностей модифікаційної мінливості. Назвіть основні константи варіаційного ряду.
6. У чому полягає комбінативна мінливість?
7. Дайте класифікацію мутаційної мінливості.
8. Дайте характеристику поліплоїдії.
9. Яка роль в еволюції органічного світу і в селекції соматичних і генеративних мутацій?
10. Назвіть причини індукованих і спонтанних мутацій. Наведіть
приклади мутагенів. 11. Дайте характеристику закону гомологічних рядів.
Селекція рослин, тварин, мікроорганізмів
Селекція (лат. selectio — добір) — наука про створення нових і поліпшення Існуючих сортів культурних рослин, порід свійських тварин і штамів мікроорганізмів, які використовує людина.
Теоретичною базою селекції є генетика та еволюційне вчення. Академік В. І. Вавилов, даючи визначення селекції як науки, зазначав її комплексність і вказував, що для успішної роботи в галузі селекції рослин потрібно враховувати вихідне сортове і видове розмаїття їхніх генотипів, роль середовища для розвитку і виявлення спадкових ознак, закономірності успадкування під час гібридизації, вплив природного добору і форми штучного добору, спрямовані на виявлення і закріплення цінних ознак.
Породи свійських тварин і сорти культурних рослин, створених людиною, є популяціями організмів. Кожний сорт рослин і кожна порода свійських тварин характеризуються своїми спадковими особливостями, морфологічними і фізіологічними ознаками, певною продуктивністю і нормою реакції на зовнішнє середовище. Кожний сорт рослин і порода тварин щонайповніше виявляють свою продуктивність лише в певних умовах, для яких вони виведені.
Порода і сорт, з одного боку, можуть включати в себе тварин і рослини з нетотожними генотипами, що дає матеріал для селекції і поліпшення цінних для господарства ознак. З іншого боку, створюють лінійні групи, які отримують самозапиленням однієї рослини або близько спорідненим схрещуванням (інбридингом) перехреснозапильних рослин, а також тварин. Лінійні групи рослин і тварин характеризуються високою гомозиготністю. Під час розведення їх у потомстві немає розщеплення і господарські ознаки зберігаються у всіх представників цієї лінії, але подальший добір у межах чистих ліній неефективний.
Завдання сучасної селекції — підвищення продуктивності сортів і порід тварин. Однак нині важливими факторами інтенсифікації рослинництва і тваринництва стають: переведення їх на промислову основу; створення короткостеблових сортів зернових культур, придатних для збирання комбайном, сортів винограду, томатів, чайних кущів, бавовнику, пристосованих до збирання врожаю машинами, сортів овочевих культур для вирощування в теплицях. У тваринництві — створення груп тварин, придатних для утримування в тваринницьких комплексах, великої рогатої худоби, придатної для машинного доїння.
У нашій країні існує розгалужена мережа селекційних установ: інститутів, селекційних станцій, сортовипробувальних ділянок, племінних господарств. У своїй роботі селекціонери використовують усю різноманітність дикорослих і культурних рослин та одомашнених тварин, бо чим різноманітніший вихідний матеріал, тим успішніше будуть реалізовані завдання, що стоять перед селекціонерами.
Велике значення мають природна мінливість і штучне отримання мутацій. Багатий матеріал дають внутрішньовидова і віддалена гібридизація.
Центри різноманітності й походження культурних рослин. Успіхи роботи селекціонерів багато в чому залежать від правильного вибору вихідного матеріалу. Методом пошуку вихідного матеріалу селекціонерів озброїв видатний генетик і селекціонер академік М. І. Вавилов. Під його керівництвом і за безпосередньої участі були організовані експедиції по всій території колишнього Радянського Союзу, Ірану, Афганістану, Середземномор'я, Абіссінії (Ефіопії), Центральної Азії, Японії, Північної, Центральної і Південної Америки. В результаті проведеної роботи М. І. Вавилов вивчив світові рослинні ресурси і встановив, що найбільшу різноманітність форм певного виду зосереджено в тих районах, де цей вид виник. На підставі вивчення зібраних матеріалів він виділив сім основних центрів походження культурних рослин: 1. Південноазій-ський тропічний (батьківщина рису, цукрової тростини, бананів, кокосової пальми тощо). 2. Східноазійський (батьківщина проса, гречки, груші, яблуні, сливи, деяких цитрусових тощо). 3. Південно-Західноазійський (батьківщина м'якої пшениці, гороху, сочевиці, бавовнику тощо). 4. Середземноморський (батьківщина маслини, буряків, капусти тощо). 5. Абіссінський (батьківщина твердої пшениці, ячменю, кавового дерева тощо). 6. Центральноамериканський (батьківщина кукурудзи, американської квасолі, гарбуза, перцю, какао, американського бавовнику тощо). 7. Південноамериканський, або Андійський (батьківщина картоплі, тютюну, арахісу, хінного дерева, ананасу тощо).
М. І. Вавилов зібрав найбільшу в світі колекцію культурних рослин, яку і нині використовують селекціонери у своїй практичній роботі. Так, відомий сорт озимої пшениці Безоста 1 був виведений П. П. Лук'яненком у результаті гібридизації взятих із колекції Вавилова аргентинських пшениць, схрещених із місцевими сортами.
Основними методами, які використовують селекціонери, є підбір, гібридизація, добір і виховання (керування Домінуванням). Гібридизація спирається на комбінативну мінливість. Завдяки їй вдається в одному гібридному організмі поєднувати цінні ознаки, які раніше існували у різних сортів рослин і порід тварин. Звичайно, для цього потрібно належним чином підібрати вихідний матеріал, а потім відібрати найкращі якості із гібридного потом-ства вихідного матеріалу.
Розрізняють дві форми добору: масовий і індивідуальний. Масовий добір грунтується на доборі за фенотипом; відбираються особини, що відповідають певному стандарту, але один і той самий фенотип може бути зумовлений різними генотипами, а один і той самий генотип може дати (в межах норми реакції) різні фенотипи, тобто слід враховувати і вплив природного добору, який завжди супроводжує штучний.
Деякі цінні ознаки виявляються лише в однієї статі, наприклад, кількість і якість молока, несучість. Визначити генотип самця за фенотипом неможливо, тому відбір лише за фенотипом не дає змоги оцінювати спадкові властивості плідників. Це, природно, уповільнює досягнення поставленої мети.
Індивідуальний добір ґрунтується на відбиранні особин з відомим генотипом за аналізом продуктивності потомства. Тому в разі індивідуального добору бажаний результат досягається швидше. Робота селекціонера у цьому разі розпочинається із створення групи особин з однаковими генотипами. Найлегше це отримати у самозапильних рослин.
Потомство однієї самозапиленої рослини, яке має однорідний гомозиготний генотип, називають чистою лінією. Однорідність генотипу у перехреснозапильних рослин і у тварин досягається схрещуванням близько споріднених форм (інбридингом). Так можна отримати групу особин з однаковим гомозиготним генотипом. Вони дістали назву інбредних ліній. Генотип чистих і інбредних ліній більш-менш стабільний, однак і в них відбуваються мутації, тому їх стабільність не абсолютна. Чисті та інбредні лінії мають злижеш життєздатність, стійкість проти захворювань, тому в селекційній практиці зазвичай створюють кілька чистих (інбредних) ліній, між якими далі проводять схрещування. Це називають міжлінійною гібридизацією, яка часом приводить до створення високопродуктивних рослин і тварин.
Гетерозис, або "гібридна сила", може спостерігатися в першому поколінні в разі гібридизації різних порід тварин і сортів рослин (зокрема, інбредних ліній). Виявляється він у підвищенні життєздатності, росту та деяких інших особливостей, у зв'язку з чим його широко використовують у сільськогосподарській практиці. Для отримання найбільшого ефекту від гетерозису в разі між лінійної гібридизації створюють кілька відмінних між собою чистих (або інбредних) ліній, потім випробовують гібридизацію між ними і встановлюють ті комбінації, які дають
найбільший економічний ефект гетерозису. Ці лінії зберігають і використовують для отримання товарного гібридного насіння.
Найбільший економічний ефект від використання гетерозису отримано для кукурудзи: підвищення врожаю зерна на 20—30 %. Ефект гетерозису добре виражений у цукрових буряків, деяких овочевих культур (цибуля, огірки, томати, баклажани, сорго) та ін. У птахівництві внаслідок схрещування несучих порід курей, наприклад леггорнів з австралорпами, род-айлендами та іншими, несучість гібридів зростає на 20—25 яєць на рік. Схрещування м'ясних порід курей поліпшує якість м'яса і прискорює темпи росту курчат. Такі гібридні курчата дістали назву бройлерів.
У свинарстві, вівчарстві, скотарстві міжпородні схрещування підвищують швидкість дозрівання, забезпечують збільшення живої маси, якості м'яса.
Ефект гетерозису можна закріпити шляхом вегетативного розмноження рослин (картопля, плодові дерева) або подвоєння числа хромосом, тобто поліплоїдизації.
Є кілька гіпотез пояснення причин гетерозису. Згідно з однією з них, у гібридів збільшується число домінантних алелів, які впливають на розвиток сприятливої ознаки. Наприклад, якщо на інтенсивність росту позитивно впливають домінантні алелі А і В відповідних генів, то в результаті схрещування представників двох рас із генотипами ААЬЬ і ааВВ гібрид АаВЬ виявиться вищим від батьківських форм. Згідно з другою гіпотезою, у багатьох алелях гетерозиготи можуть мати більш виражені ознаки, ніж домінантні гомозиготи. Ймовірно, після детальнішого дослідження явища гетерозису обидві гіпотези виявляться взаємодоповиювальними.
Поліплоїдія. Важливу роль у створенні нових сортів рослин відіграє поліплоїдія. Поліплоїдні форми трапляються у багатьох рослин. Не виключено, що еволюція деяких груп квіткових рослин ішла шляхом поліплоїдизації. Більшість культурних рослин — поліплоїдні, вони економічно доцільніші за диплоїдні. У поліплоїдних рослин клітини зазвичай мають більші розміри і вся рослина більша за диплоїдну. Поліплоїди часто стійкіші до несприятливих факторів середовища. На півночі і в горах багато видів рослин — поліплоїдні. В експериментах і в селекційній практиці, щоб отримати поліплоїдні рослини, на них діють різними факторами зовнішнього середови-Ща: радіоактивним випромінюванням, хімічними речовинами, критичною температурою. Наприклад, для отримання поліплоїдів досить зручним виявився колхіцин, оскільки він під час мітозу руйнує веретено поділу, але не перешкоджає подвоєнню хромосом.
Селекціонери для отримання поліплоїдних форм часто використовують мутагени. У нашій країні останнім часом великого поширення набули високоурожайні полі-плоїдні сорти гречки, цукрових буряків, проса, кукурудзи, льону, редиски та деяких інших рослин.
Віддалена гібридизація відіграє істотну роль у селекції. Зазвичай схрещують сорти, які належать до одного виду, але іноді проводять схрещування міжвидове і міжродове.. Виведено перспективні гібриди пшениці і пирію, жита і пшениці, пшениці і ячменю (тритикале) та ін.
Основна складність створення стабільних гібридних форм спричинена безплідністю більшості отриманих гібридів. Причина безплідності полягає в тому, що батьківські негомологічні хромосоми під час мейозу не здатні кон'югувати, а отже, у гібридів не можуть утворюватися гамети.
Вихід із цього становища знайшов генетик Г. Д. Карпеченко. Він отримав гібрид редиски і капусти. Кожний із вихідних батьківських видів мав по 18 хромосом у дип-лоїдному наборі. Спершу хромосомний набір гібрида складався також із 18 хромосом, куди входив гаплоїдний набір хромосом капусти (9 хромосом) І гаплоїдний набір хромосом редиски (9 хромосом). Оскільки хромосоми вихідних батьківських видів були не гомологічні, то кон'югація між ними не відбувалася, мейоз не здійснювався, статеві клітини не утворювалися. Гібрид був безплідним. Г. Д. Карпеченко домігся подвоєння числа хромосом (тобто отримав тетраплоїд). Тепер у клітинах гібрида був диплоїдний набір хромосом капусти і диплоїдний набір хромосом редиски. Мейоз став можливим. Гібрид виявився здатним до розмноження. Метод отримання поліплоїдних міжвидових гібридів значно поширився у світовій селекційній практиці.
Методи роботи І. В. Мічуріна. У своїй роботі ї. В. Мічурін широко застосовував гібридизацію. При цьому він враховував складну природу гібридів. Він вважав, що гібридні сіянці на тих чи інших стадіях розвитку проходять критичні періоди, під час яких і відбувається реалізація різноякісних батьківських генів. Грунтуючись на цьому припущенні, І, В. Мічурін застосовував прийоми виховання сіянців, впливаючи на них факторами середовища. Він використовував різні способи обробітку грунту, внесення добрив, метод ментора. Метод ментора полягає в тому, що вкрону молодого гібридного сіянця (підщепи) шеплять живець від того сорту, властивості якого бажають мати в гібрида. Іноді гібрид щеплять на відповідну прищепу.
Результативність менторального впливу залежала від тривалості дії ментора, співвідношення віку ментора і гібрида та від ступеня розвитку їхнього листя. Чим старіший ментор, чим більша його крона порівняно з гібридною прищепою та чим триваліша дія, тим кращий результат.
І. В. Мічурін широко використовував також і віддалену гібридизацію. Він встановив, що в разі гібридизації південних сортів з місцевими зазвичай домінують ознаки місцевих сортів. Щоб уникнути цього, він підбирав вихідні форми з різних географічних районів, тобто схрещував географічно віддалені форми. Так, сорт груші Бере зимова він вивів гібридизацією південного французького сорту Бере Рояль з дикою уссурійською грушею. За такої гібридизації у гібрида спадкова основа обох вихідних батьківських форм виявилася в незвичних природних умовах. Змінюючи умови вирощування гібридів, І. В. Мічурін виховував у них господарсько цінні ознаки, взяті як від одного, так і від другого з батьків. У цьому конкретному випадку сорт груші Бере зимова успадкував від південного батька великі плоди, високі смакові якості і здатність до тривалого зберігання, а від уссурійської груші — морозостійкість.
У зв'язку з тим, що організми під впливом середовища здатні змінюватися в межах норми реакції, зовнішніми факторами можна впливати на фенотиповий прояв ознак. Цим скористався І. В. Мічурін під час вирощування гібридів плодових дерев, змінюючи певні умови на різних стадіях онтогенезу. Домінування ознак, які в навколишньому середовищі зустрічають сприятливіші умови для свого розвитку, І. В. Мічурін назвав керуванням домінування.
Щоб подолати несхрещуваність у разі віддаленої гібридизації, І. В. Мічурін розробив низку методів.
Метод вегетативного зближення полягає в попередньому щепленні одного виду рослин на інший. Внаслідок цього змінюється хімічний склад тканин, що, мабуть, сприяє проростанню пилкових трубок в маточці материнської рос-лини. Так можна домогтися запліднення під час гібриди-зації таких видів, які зазвичай не схрещуються. Саме за цим методом було отримано гібриди груші і горобини, яблуні і груші, вишні і японської черемхи (церападус), айви і груші.
Метод посередника полягає ось у чому: якщо схрещу, вання між двома віддаленими формами А і В не вдається, то підшукують третю С, яка схрещується з однією з перших двох (наприклад, з А). Отриманий від цього схрещування гібрид D, що має розхитану спадковість, порівняно легко схрещується з другою з двох початково вибраних для гібридизації форм (з В). Гібрид D і є "посередником", сполучною ланкою між А і В. Цим методом І. В. Мічурін вивів сорт північного персика.
Метод суміші пилку реалізується використанням для штучного запилення суміші пилку кількох сортів батьківського виду і материнської рослини, а іноді ще й з додаванням пилку інших видів рослин. Суміш складають у певних кількісних поєднаннях, що створює найсприятливіші умови для вибіркового запліднення, тобто серед безлічі пилкових клітин виявляються найбільш біологічно відповідні жіночій гаметі і запліднення відбувається. Більшість виведених І. В. Мічуріним сортів були складними гетерозиготами. Щоб зберегти їхні якості, їх слід розмножувати вегетативно: відсадками, щепленням тощо, бо в разі статевого розмноження вони розщеплюватимуться.
І. В. Мічурін багато уваги приділяв акліматизації південних плодових рослин, започаткував просування на північ винограду, абрикоса, черешні тощо.
Видатні заслуги у виведенні нових сортів сільськогосподарських рослин М. В, Цицина, П. П. Лук'яненка, В. М. Ремесла, Ф. Г. Кириченка, В. Є. Писарєва, В. С. Пустовойта, І. М. Хаджинова та інших селекціонерів.
М. В. Цицин розробив теоретичні основи створення нових видів і форм рослин віддаленою гібридизацією культурних і декоративних рослин. Він вивів пшенично-пирійні, пшеничноелімусні, житньо-пирійні та інші гібриди, які є практично новими видами рослин.
Значним досягненням селекціонерів є сорти озимих пшениць Безоста, Аврора, Кавказ, які вивів П. П. Лук'яненко. В них сконцентровані цінні властивості — порівняно коротка і міцна солома, висока врожайність, зимостійкість, високі якості борошна. Нині ці сорти у світовому землеробстві посідають перше місце за площею посівів — понад 11 млн га.
В. М. Ремесло вивів ряд сортів озимої пшениці: Миро-нівська 264, Миронівська ювілейна, Миронівська 807,
Іллічівка та інші, площі під якими становлять понад 11 % площ світових озимих пшениць. На їх основі створено близько 40 нових сортів пшениць. Деякі з них можуть давати врожаї до 100 ц/га. Сорт Миронівська яра також має високу врожайність — 45—64 ц/га.
ф, Г. Кириченко вивів ряд зимостійких сортів озимої м'якої пшениці, таких як Одеська 3, Одеська 12, Одеська 16, Одеська 26, Степова. Вперше створено озиму тверду пшеницю, найурожайнішими сортами якої с Мічурінка, Ново-мічурінка, Одеська ювілейна.
В. Є. Писарєв вивів нові високоврожайні сорти пшениці- Методом генної інженерії отримано новий сорт пшениці Киянка з високими врожайністю і стійкістю проти захворювань.
І. М. Хаджинов досяг великих успіхів у селекції кукурудзи. Він одним із перших відкрив явище цитоплазматичної чоловічої стерильності у кукурудзи і використав її для поліпшення насінництва цієї культури. Він вивів понад 20 високопродуктивних гібридів кукурудзи, із них 14 — подвійних міжлінійних гібридів.
Т. Є. Тарасенко та І. Д. Прохожай вивели високоврожайні сорти ярового ячменю: Донецький 4 та Донецький 6, які райо-новано в багатьох областях України.
Заслуженою популярністю користуються сорти картоплі, які вивела Н. А. Лебедєва.
Кілька поліплоїдних сортів буряків отримали М. П. Ду-бінін, А. Н. Лутков, В. О. Панін.
В. С. Пустовойт створив понад 20 високоолійних, стійких проти хвороб сортів соняшнику. Вміст жиру в абсолютно сухому насінні становить понад 55 % (вміст у вихідних формах не перевищував 35 %).
Селекція тварин. Усі свійські тварини походять від диких предків. Найраніше, у середині кам'яного віку був одомашнений собака; предками його були вовк і, можливо, шакал. Наприкінці кам'яного віку були одомашнені свині, вівці, кози, велика рогата худоба, згодом — коні. Свині походять від диких європейських і азійських кабанів, вівці — від диких європейських овець, кози — від гвинторогого козла, велика рогата худоба — від тура, коні — від тарпана і коня Пржевальського. З глибокої давнини людина вела безперервний добір свійських тварин, залишаючи на розплід як плідників ті особини, в яких та чи інша спадкова ознака була найпомітнішою і найбільше задовольняла вимогам (економічним, естетичним тощо). Усі інші особини не допускалися до розмноження і використовувалися для задоволення господарських потреб (на м'ясо, як робоча худоба тощо). Спочатку цей добір був несвідомим (для задоволення потреб, без мети поліпшення чи виведення), а далі поступово почав набувати характеру примітивного методичного добору.
Завдяки добору упродовж тисячоліть сформувалися численні місцеві породи, пристосовані до специфічних умов різних територій проживання людини та її потреб. Нині для виведення нових і поліпшення існуючих порід свійських тварин селекціонери використовують у принципі такі самі методи, як і в рослинництві.
Велике значення має добір плідників за цінними господарськими ознаками та екстер'єром тварин. Екстер'єр — сукупність фенотипових ознак тварини. Беруться до уваги будова тіла і співвідношення розмірів частин тіла, оскільки зовнішні форми тварини та її внутрішні якості взаємозв'язані. За екстер'єром визначають породність тварин, типовість, індивідуальні особливості, вікову мінливість, кондицію, стан здоров'я і тип продуктивності.
У разі селекції коней, свиней, овець, великої рогатої худоби м'ясних порід плідників оцінюють за фенотипом (екстер'єром) і за якістю потомства від них. У разі селекції великої рогатої худоби молочних порід добір проводять у три етапи. Попередній добір самців грунтується на відомостях про молочність матерів і сестер та на ознаках екстер'єру. Далі оцінюють плідників за продуктивністю потомства. Нарешті, кращих плідників схрещують з дочками, щоб з'ясувати, чи не мають вони летальних та інших небажаних генів. Від доброго самця можна дістати велике потомство, особливо якщо застосовувати штучне запліднення.
Велика заслуга щодо виведення нових порід свійських тварин належить видатному селекціонеру М. Ф. Іванову, який вивів високопродуктивні породи свиней і овець. Робота проводилась в Асканії-Нова.
Завезені в Україну високопродуктивні білі англійські свині виявилися непристосованими до місцевих умов. І навпаки, місцева безпородна свиня характеризувалася витривалістю, доброю плодючістю, невибагливістю, але мала погані м'ясні якості. Селекціонер провів схрещування кнура білої англійської породи із свиноматками місцевої породи. Гібридні самки першого покоління знову були схрещені з чистопородним кнуром білої англійської породи. Із потомства були відібрані плідники з найціннішими ознаками, добре пристосовані до місцевих умов. Від них він
початку отримав кілька інбредних ліній тварин, у результаті схрещування між якими вивів нову породу свиней — українську степову білу.
Користуючись цим самим методом, М. Ф. Іванов вивів нову породу овець — асканійську тонкорунну.
Віддалена гібридизація свійських тварин. Схрещування свійських тварин з дикими предками дає плодюче потомство і може бути використане з метою селекції. М. Ф. Іванов у результаті схрещування тонкорунних овець з одним із підвидів диких баранів (муфлоном) вивів нову породу гірського мериноса. Казахський архаромеринос також виведений у результаті схрещування тонкорунних овець з диким бараном (архаром). Внаслідок схрещування великої рогатої худоби з горбатою худобою (зебу) отримано цінні групи молочної худоби.
Здебільшого міжвидові гібриди безплідні, оскільки у них не відбувається мейоз. Проте таке потомство може мати дуже велике практичне значення, оскільки в нього добре виражений гетерозис. Гібриди кобили з ослом — мули — дуже витривалі, мають велику фізичну силу, за тривалістю життя значно перевищують батьківські види; гібриди яка з великою рогатою худобою переважають вихідні форми за масою і здатністю до відгодівлі. У гібридів яка і великої рогатої худоби безплідні самці, а самки — плодючі. Гібриди одногорбого і двогорбого верблюдів також переважають вихідні види за розмірами і витривалістю. Тому для отримання таких гібридів з давніх часів проводились міжвидові схрещування.
Селекція відіграє певну роль у збереженні різноманітності органічного світу. Коли на початку XX ст. в Європі збереглися лише поодинокі особини зубрів, то для врятування виду було проведено схрещування зубрів з бізонами. Нині, можливо, в природі вже зник кінь Пржевальського. Збереглося кілька груп цих тварин у зоопарках і заповіднику АсканіяНова. Для врятування виду і збереження гетерозиготності зоопарки різних країн обмінюються окремими особинами. Проведена гібридизація із свійськими кіньми і гібридів — з дикими кіньми. Частина коней Пржевальського відправлена на батьківщину, в Монголію, для відновлення зниклої популяції.
Селекція мікроорганізмів. Чимало галузей харчової промисловості пов'язано з діяльністю мікроорганізмів (випікання хліба, виноробство, пивоваріння, отримання спирту, кисломолочних продуктів тощо). Лікарські засоби — антибіотики — також отримують у результаті діяльності
мікроорганізмів. Для вдосконалення їхньої продуктивності використовують методи селекції. За допомогою рентгенів. випромінювання і хімічних речовин прискорюють мутагенний процес і добором створюють кращі раси (штами) мікроорганізмів. Цим способом у селекційних рас у тисячі разів вдалося підвищити вихід низки антибіотиків, зокрема пеніциліну, порівняно з вихідними штамами мікроорганізмів, взятими для селекції з природи. Отримано дріжджові гриби, які синтезують кормовий білок із парафінів нафти, природного газу, відходів рослинництва (соломи зернових культур, стебел соняшнику, відходів лісового господарства). У кондитерській промисловості широко використовують лимонну кислоту, яку отримують в результаті життєдіяльності спеціально виведених мікроорганізмів. У світі нині виробляють близько 400 тис. т цього продукту. Таку кількість його не змогли б забезпечити жодні цитрусові плантації.
Створюються штами мікроорганізмів, які здатні вилучати цінні метали із руд, промислових відходів для виробництва бактеріальних добрив, стимуляторів росту і мікробіологічних засобів захисту рослин від шкідників і хвороб. Для виробництва цих та інших речовин за допомогою мікроорганізмів створено спеціальний напрям народного господарства — мікробіологічну промисловість. У промислових масштабах нині виробляють багато амінокислот, які використовують як кормові добавки, вітамінів тощо.
Основні напрями біотехнології. Біотехнологією називають свідоме виробництво потрібних людині продуктів і матеріалів за допомогою біологічних об'єктів і процесів. Біотехнологія виникла на зорі цивілізації, коли первісна людина навчилася не просто збирати корисні для себе рослини, а й вирощувати їх на оброблюваних полях, не тільки полювати на диких звірів, а й розводити приручених тварин. Ця первісна біотехнологія значно полегшила життя людини, бо вона почала отримувати значно більше їжі і сировини для виготовлення одягу, а затрачала на це менше праці. Поступово біотехнологія вдосконалювалася, людина почала створювати нові сорти культурних рослин і породи свійських тварин, учитись ефективніше їх використовувати.
На початку XX ст., коли, здавалося б, людина навчилася отримувати від природи все, що можна, виникла нова наука — генетика. Проте минуло майже 50 років, доки її результати почали приносити користь. Свідомим поєднанням випадкових спадкових змін (мутацій) людина навчилася створювати все досконаліші сорти і породи, а також різновиди (штами) корисних мікроорганізмів, на основі яких виникла мікробіологічна промисловість (див. вище).
У 50-х роках нашого століття виникла нова наука — молекулярна біологія, а ще через 20 років на її основі — генна інженерія. Всього 10 років знадобилось їй, щоб дати біотехнології нову зброю: принципову можливість свідомо створювати організми, які б продукували сполуки і здійснювали процеси, необхідні людині. Тобто можна стверджувати, що виникла нова біотехнологія, яка обіцяє людині небувалий прогрес.
Для досягнення мети в генній інженерії використовують такі способи; злиття соматичних клітин або протопластів різних клітин одного виду чи навіть різних видів організмів (соматична гібридизація), перенесення ядер із клітини в клітину, хромосом або їхніх фрагментів {клітинна інженерія) чи окремих генів (генна інженерія). Суть генної інженерії полягає у штучному створенні (хімічний синтез, перекомбінації відомих структур) генів з конкретними потрібними людині властивостями і введення його у відповідну клітину (нині це найчастіше бактеріальні клітини, наприклад кишкова паличка) — створення "штучної" бактерії-лабораторії для вироблення необхідного для людини продукту.
Методи генної інженерії дали змогу синтезувати деякі білки у промислових масштабах. Поки що це виявилось можливим і економічно доцільним лише для кількох білків людини, які використовуються у медичній практиці і є видоспецифічними, тобто не можуть бути замінені на аналогічні білки тварин. Йдеться насамперед про інсулін, інтерферон і гормон росту — соматотропін. Список цей розширюється.
Інсулін необхідний для лікування діабету, соматотропний гормон — природний стимулятор росту, інтерферон — білкова речовина, яка сприяє активній боротьбі клітин організму з вірусами.
Контрольні запитання і завдання
1. Дайте визначення селекції та назвіть її завдання.
2. Що називають сортом культурних рослин і породою свійських тварин?
3. Які дослідження провів М. І. Вавилов щодо визначення походження культурних рослин?
4. Чи впливає природний добір на виведення сортів рослин і порід тварин?
5. Які методи використовують у селекційнііі роботі?
6. Що таке чисті лінії і міжлішйна гібридизація?
7. Що таке гетерозис І яка його роль у рослинництві і тваринництві?
8. Яке значення в селекції рослин має поліплоїдія?
9. Яка роль віддаленої гібридизації в селекції рослин і тварин? Чим пояснюють безплідність більшості віддалених гібридів? Чи можна її подолати?
10. Якими методами користувався І. В. Мічурін для виведеная нових сортів?
11. Дайте характеристику досягнень вітчизняних селекціонерів.
12. У чому полягають особливості селекції тварин?
13. Перелічіть напрями біотехнології.