

ЗМІСТ
Автори: А. О. Слюсарєв, О. В. Самсонов, В. М. Мухін, Є. Є. Федосова, А. П. Щеулов, В. П. Сисмєєв, Т. І. Самойленко
Редакція літератури з природничих наук Редактори: Я. А. Серебрякова, Л. Є. Канівєць
Біологія: Навч. посіб. / А. О. Слюсарєв, О. В. Сам-Б63 сонов, В. М. Мухін та ін.; За ред. та пер. з рос. В. О. Мотузного. — 3-тє вид., випр. і допов. — К.: Вища шк., 2002. — 622 с: іл.
І8ВИ 966-642-027-9
Викладено основні положення загальної біології, ботаніки, зоології, анатомії та фізіології людини. Третє видання (2-ге вид. — 1995 р.) доповнене деякими відомостями відповідно до змін у навчальній програмі.
Для учнів спеціалізованих шкіл, ліцеїв та гімназій.
ІЗВИ 966-642-027-9
УДК 57(075.8) ББК 28я729
© А. О. Слюсарєв, О. В. Самсонов,
В. М. Мухін та ін., 1995 © А. О. Слгосарєв, О. В. Самсонов,
В. М. Мухін та ін., 2001, зі змінами © Переклад на українську мову
В. О. Мотузного, 1991
В с т у п (А О. Слюсарєв)................................................. 6
ЗАГАЛЬНА БІОЛОГІЯ
Клітинні і неклітинні форми життя (А П. Щеулов).......... 10
Будова і функції клітини (А П. Щеулов)......................... 13
Хімічна організація клітини (А П. Щеулов).................... 28
Обмін речовин і перетворення енергії в клітині(А П. Щеулов).. 39
Розмноження та, індивідуальний розвиток організмів
(А П. Щеулов)............................................................... 52
Основи генетики і селекції (А О. Слюсарєв)..................... 76
Основні закономірності спадковості.............................. 76
Закономірності мінливості.......................................... 101
Селекція рослин, тварин, мікроорганізмів..................... 113
Еволюційне вчення (А О. Слюсарєв).............................. . 126
Загальна характеристика біології в до дарвінівський період ..... 126
Дарвінізм........................................................... ....... 134
Закономірності розвитку органічного світу.................. 163
Розвиток життя на Землі .............................................. 172
Походження людини (А П. Щеулов) ............................ .. 183
Основи екології (А О. Слюсарєв)................................. ... 195
Біосфера і людина (А О. Слюсарєв)........................... ..... 211
БОТАНІКА
Будова рослин (Є. Є. Федосова)................................... ..... 225
Рослинні тканини...................................................... . 225
Вегетативні органи.................................................. .... 232
Квітка, плід, насіння............................,...................... 263
Рослина — цілісний організм........................................ . 282
Бактерії (О. В. Самсонов) ............................................... 285
Водорості (Є. Є. Федосова).............................................. 291
Царство Гриби (О. В. Самсонов)..................................... 300
Лишайники (Є. Є. Федосова).......................................... 309
Підцарство Вищі рослини (Т. І. Самойленко).................. 314
Відділ Мохоподібні, або Бріофіти................................. 315
Відділ Папоротеподібні, або Поліподіофіти................... 319
Відділ Хвощоподібні, або Еквізетофіти.......................... 322
Відділ Плауноподібні, або Лікоподіофіти...................... 323
Відділ Голонасінні, або Пінофіти (Є. Є. Федосова)......... .. 324
Відділ Покритонасінні, або Магноліофіти
(Є. Є. Федосова).......................................................... 329
ЗООЛОГІЯ
Тип Найпростіші (О. В. Самсонов).................................. 341
Клас Саркодові ......................................................... . 343
Клас Джгутикові........................................................ 345
Клас Споровики ......................................................... 347
Клас Інфузорії ........................................................... 349
Тип Кишковопорожнинні (О. В. Самсонов)...................... 352
Клас Гідрозої............................................................. 353
Клас Сцифомедузи ..................................................... 356
Клас Коралові поліпи................................................. 358
Тип Плоскі черви (О. В. Самсонов)................................ . 359
Клас Війчасті черви ................................................... 360
Клас Стьожкові черви................................................. 362
Клас Сисуни, або Трематоди ........................................ 365
Тип Первиннопорожнинні, або Круглі черви (О. В. Самсонов)... 367
Клас Круглі черви, або Нематоди............................ ..... 368
Тип Кільчасті черви, або Кільчаки (О. В. Самсонов, А. О. Слюсарев)............................................................ . 374
Тип Молюски, або М'якуни (О. В. Самсонов) ................... 380
Клас Черевоногі......................................................... 381
Клас Двостулкові........................................................ 382
Клас Головоногі ......................................................... 385
Тип Членистоногі (О. В. Самсонов, А. О. Слюсарєв)....... . 386
Клас Ракоподібні........................................................ 389
Клас Павукоподібні.................................................... 394
Клас Комахи.............................................................. 398
Тип Хордові (О. В. Самсонов, А. О. Слюсарєв).................. 415
Клас Ланцетники....................................................... 416
Надклас Риби ............................................................ 419
Клас Земноводні......................................................... 430
Клас Плазуни ............................................................ 437
Клас Птахи................................................................. 443
Клас Ссавці ............................................................... 456
ЛЮДИНА ТА її ЗДОРОВ'Я
Анатомія, фізіологія і гігієна людини (В. П. Сисмєєв)...... 469
Гігієнічні аспекти охорони навколишнього середовища
(В. 77. Сисмєєв)............................................................. 470
Загальний огляд організму людини (В. П. Сисмєєв)......... 471
4
Опорно-рухова система (В. П. Сисмєєв)............................ 478
Огляд м'язової системи людини (В. П. Сисмєєв).............. 491
Кров (В. П. Сисмєєв) ..................................................... 495
Кровообіг (В. П. Сисмєєв)............................................... 502
Дихання (В. М. Мухін) ................................................. 509
Травлення (В. М. Мухін)............................................... 519
Вітаміни (В. М. Мухін) ................................................. 534
Виділення (В. М. Мухін)............................................... 538
Шкіра (В. М. Мухін)..................................................... 541
Нервова система (В. М. Мухін) ...................................... 547
Вища нервова діяльність (В. М. Мухін).......................... 561
Аналізатори. Органи чуття та їх значення (В. М. Мухін) 581
Залози внутрішньої секреції (В. М. Мухін) .................... 594
Розвиток організму людини (В. П. Сисмєєв)................... 599
Додаток (А Я. Щеулов)............................................ 605
Предметний покажчик.................................. 609
Біологія — наука про життя. Нині — це комплекс наук про живу природу. Об'єктом вивчення біології є живі організми — віруси, бактерії, водорості, гриби, лишайники, вищі рослини та тварини (від найпростіших одноклітинних організмів до людини), їх різноманітність, будова тіла й органів, розвиток, поширення, еволюція та форми співіснування в екологічних системах.
Перші відомості про живі організми почала нагромаджувати ще первісна людина. Живі організми давали їй їжу, матеріал для одягу і житла. Вже тоді людина не могла обійтися без знань про властивості рослин, місця їх зростання, строки дозрівання насіння і плодів, місця мешкання і звички тварин, на яких вона полювала, хижих і отруйних тварин, які могли загрожувати її життю. Так поступово накопичувалися відомості про живі організми.
Приручення тварин та спроби вирощування рослин потребували глибших відомостей про живі істоти. Значний фактичний матеріал про живі організми зібрав відомий лікар Давньої Греції Гіппократ (близько 460—377 р. до н. є.). Це були відомості про будову тіла тварин і людини, кістки, м'язи, сухожилки, головний і спинний мозок. Перша велика праця із зоології належить давньогрецькому філософу Аристотелю (384—322 р. до н. є.), який описав понад 500 видів тварин. Аристотель цікавився будовою тіла і способом життя тварин, започаткував науку зоологію. Першу спробу систематизувати знання про рослини зробив давньогрецький природодослідник Теофраст (372—287 р. до н. є.). Розширенням знань про будову людського тіла наука завдячує римському лікарю і природодосліднику Галену (близько 130—200 р. к. є.), який досліджував будову організму тварин, розтинаючи трупи мавп. Результати його праці впливали на розвиток природознавства і медицини упродовж кількох століть.
В епоху середньовіччя під гнітом церкви наука розвивалася дуже повільно. Важливим рубежем у розвитку науки стала епоха Відродження. Вже у XVIII ст. почали розвиватися як самостійні науки ботаніка, зоологія, анатомія людини, фізіологія.
Поступово нагромаджувалися відомості про різноманітність видів, будову тіла тварин і людини, індивідуальний розвиток, функції органів рослин і тварин. Упродовж багатовікової історії розвитку біології найвизначнішими віхами у вивченні органічного світу можна вважати впровадження принципів систематики, запропонованих К. Ліннеєм; винайдення мікроскопа; створення Т. Шванном клітинної теорії; створення Ч. Дарвіном еволюційної теорії; відкриття Г. Менделем основних закономірностей спадковості; використання електронної мікроскопії для біологічних досліджень; розшифрування генетичного коду; створення вчення про біосферу.
Нині науці відомо близько 1 млн 500 тис. видів тварин і понад 500 тис. видів рослин. Вивчення розмаїття рослин і тварин, особливостей будови їхніх організмів та життєдіяльності має велике значення для з'ясування їх ролі в існуванні біосфери. Біологічні науки — основа для розвитку рослинництва, тваринництва, медицини, біотехнології.
Найдавніші біологічні науки — анатомія і фізіологія людини — становлять теоретичний фундамент медицини. Кожна людина повинна мати уявлення про будову і функції власного організму, щоб у разі потреби зуміти надати першу допомогу, свідомо оберігати своє здоров'я і дотримуватись правил гігієни.
Упродовж століть ботаніка, зоологія, анатомія і фізіологія розвивалися як самостійні, ізольовані науки. Лише в XIX ст. було виявлено закономірності, спільні для всіх живих істот. Так виникли науки, що вивчають загальні закономірності життя. До них належать: цитологія — наука про клітину; генетика — наука про мінливість і спадковість; екологія — наука про взаємозв'язки організмів із середовищем і між собою у природних угрупованнях тощо. У курсі загальноосвітньої школи основи цих наук є предметом вивчення загальної біології.
Пропонована читачам книга рекомендована для учнів спеціалізованих шкіл, ліцеїв та гімназій біологічного спрямування. Нею також можуть скористатися вступники до університетів, сільськогосподарських, лісотехнічних, медичних та педагогічних вищих навчальних закладів. Посібник написано відповідно до навчальної програми для учнів старших класів загальноосвітньої школи.
Деякі програмні матеріали у посібнику подано конспективно, бо вони легко засвоюються учнями всіх рівнів навчання, інші — дещо ширше, що важливо для створення повнішого уявлення про той чи інший розділ біології. Виклад матеріалу розпочинається з розділу "Загальна біологія" для формування загально біологічних понять на ранніх етапах вивчення цього предмета. Саме тому спочатку розглядаються будова і функції клітин, потім явища розмноження, індивідуального розвитку, спадковості та мінливості. Завершується розділ відомостями з еволюційного вчення та екології. Саме така послідовність дасть змогу творчо, з розумінням підійти до вивчення наступних розділів ("Ботаніка", "Зоологія", "Людина та її здоров'я"), свідомо використовувати загально-біологічні уявлення про закономірності життя представників різних царств живої природи. У посібнику поряд Із загальновживаними поняттями в міру можливості використано і наукову термінологію, що полегшує перехід від вивчення шкільного курсу до подальшого поглибленого засвоєння матеріалу у вищому навчальному закладі.
Посібник ілюстровано. Малюнки значно поліпшать засвоєння матеріалу. У процесі вивчення окремих розділів біології потрібно навчитися розв'язувати задачі, тому в додатку вміщено приклади розв'язування типових задач з молекулярної біології та генетики. Наприкінці кожного розділу наведено запитання для самоконтролю рівня засвоєння матеріалу.
Під час складання цього посібника авторський колектив — співробітники кафедри біології Донецького медичного університету та редактор і перекладач, доцент кафедри фізіології людини і тварин Київського національного університету імені Тараса Шевченка — використали свій багаторічний досвід викладання цього предмета на підготовчих відділеннях і приймання вступних іспитів.

КЛІТИННІ І НЕКЛІТИННІ ФОРМИ ЖИТТЯ
Основними формами життя на Землі є організми клітинної будови. Цей тип організації характерний для всіх видів живих істот, за винятком вірусів, які розглядають як неклітинні форми життя. Вони настільки малі, що лише в кілька разів перевищують розміри великих молекул білків. Віруси мають розміри 10—275 нм. їх можна побачити лише під електронним мікроскопом. Вони легко проходять крізь пори спеціальних фільтрів, що затримують усі бактерії і клітини багатоклітинних організмів. Віруси були відкриті у 1892 р. російським фізіологом рослин і мікробіологом Д. І. Іванівським під час вивчення хвороби тютюну.
Віруси — збудники багатьох хвороб рослин і тварин. Вірусними хворобами людини є кір, грип, гепатит А (хвороба Боткіна), поліомієліт (дитячий параліч), сказ, віспа тощо.
Під електронним мікроскопом різні види вірусів мають форму паличок або кульок. Окрема вірусна часточка складається з молекули нуклеїнової кислоти (ДНК або РНК), скрученої в клубок, і молекул білка, розміщених у вигляді своєрідної оболонки навколо молекули кислоти (капсид). Віруси не здатні самостійно синтезувати нуклеїнові кислоти і білки, з яких вони складаються. Розмноження вірусів можливе лише в разі використання ферментних систем клітин. Потрапивши у клітину, віруси змінюють і перебудовують обмін речовин у ній, внаслідок чого клітина починає синтезувати молекули нових вірусних часточок. Поза клітинами віруси переходять у кристалічний стан, що сприяє їх збереженню.
У житті вірусів можна виділити такі етапи: прикріплення вірусу до клітини, вторгнення вірусу в клітину, латентну стадію, утворення нового покоління вірусів, вихід вібріонів. У латентну стадію вірус ніби зникає. Його не вдається виявити або виділити з клітини, але в цей період уся клітина синтезує необхідні для вірусу білки і нуклеїнові кислоти, в результаті чого утворюється нове покоління вібріонів.
Проникнення вірусу в клітину організму хазяїна розпочинається із взаємодії вірусної часточки з поверхнею клітини, на якій є особливі рецепторні ділянки. Оболонка часточки вірусу має відповідні прикріпні білки, які "впізнають" ці ділянки. Саме цим зумовлена висока специфічність вірусів стосовно клітин-хазяїв: часто віруси уражують лише певний тип клітин якогось виду організмів. Так, вірус поліомієліту уражує лише нервові клітини людини, а вірус тютюнової мозаїки — клітини листків тютюну. Якщо часточка вірусу прикріплюється не до рецепторних ділянок, а до інших місць на поверхні клітини-хазяїна, то зараження останньої може і не відбутися. Отже, наявність рецепторних ділянок на поверхні клітини визначає її чутливість до того чи іншого виду вірусів.
Усередину клітини-хазяїна вірус може проникнути різними шляхами. Часом оболонки вірусних часточок зливаються з клітинною мембраною (як у вірусу грипу), і ДНК виявляється у цитоплазмі клітини, іноді вірусна часточка потрапляє в клітину шляхом піноцитозу, після чого ферменти клітини-хазяїна розщеплюють її оболонку і вивільняють нуклеїнову кислоту (вірус поліомієліту тварин). У рослинні клітини віруси можуть проникати крізь пошкоджені ділянки клітинної стінки.
У 1917 р. французький вчений Ф. д'Ерелл відкрив віруси бактерій — бактеріофаги (або фаги). Під електронним мікроскопом вони мають форму коми або тенісної ракетки розміром близько 5 нм. Коли часточка фата прикріплюється своїм тонким відростком до бактеріальної клітини, його ДНК проникає в клітину і викликає синтез нових молекул ДНК і білка бактеріофага. Через ЗО—60 хв бактеріальна клітина руйнується і з неї виходять сотні нових часточок фага, здатних спричинити зараження інших бактеріальних клітин.
Спочатку вважали, що бактеріофаги можна використовувати для боротьби з хвороботворними бактеріями. Проте згодом виявилося, що фаги швидко руйнують бактерії в пробірці, але неефективні в живому організмі. У зв'язку з цим їх використовують в основному для діагностики захворювань, виявлення бактерій.
Клітинна теорія. Відкриття і вивчення клітини стало можливим завдяки винайденню мікроскопа та вдосконаленню методів мікроскопічних досліджень. Перший опис клітини зроблено у 1665 р. англійським ученим Р. Гуком. Згодом з'ясувалося, що він відкрив не клітини (у власному розумінні терміна), а лише зовнішні оболонки рослинних клітин.
Прогрес у вивченні клітини пов'язаний з розвитком мікроскопії у XIX ст. На той час уявлення про будову клітини змінилось: за основу клітини бралася не клітинна оболонка, а її вміст — протоплазма. У протоплазмі було відкрито постійний компонент клітини — ядро. Численні відомості про тонку будову та розвиток тканин і клітин дали змогу зробити узагальнення. Таке узагальнення запропонував у 1839 р. німецький біолог Т. Шванн у вигляді сформульованої ним клітинної теорії. Він стверджував, що клітини рослин і тварин принципово подібні між собою (мал. 1).
Крім Т. Шванна та М. Шлейдена, співавторами клітинної теорії вважають німецького вченого Рудольфа Вірхова та естонського вченого Карла Бера. Р. Вірхов довів, що клітини утворюються з інших клітин внаслідок їх поділу. К. Бер відкрив яйцеклітину птахів і ссавців і довів, що
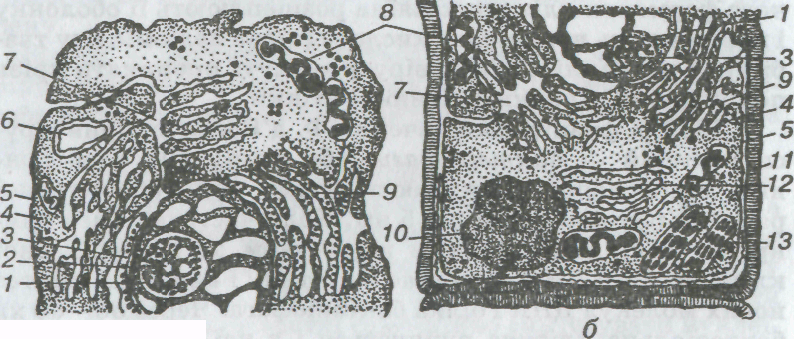
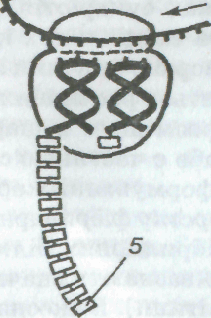
Мал. 1. Будова тваринної (а) і рослинної (б) клітин:
1 — ядро; 2 — ядерна мембрана; 3 — ядерце; 4 — протоплазма; 5 — плазматична мембрана; 6 — лізосома; 7 — комплекс Гольджі; 8 — мітохондрії; 9,12 — відповідно шорстка і гладенька ендоплазматична сітка; 10 — вакуоля; 11 — клітинна оболонка; 1З — хлоропласт
багатоклітинні організми цих тварин розвиваються з однієї клітини — заплідненої яйцеклітини (зиготи). Отже, клітина є одиницею не лише будови, а й розвитку організму.
Створення клітинної теорії стало найважливішою подією в біології, одним із вирішальних доказів єдності всієї живої природи. Клітинна теорія сприяла розвитку ембріології, гістології і фізіології. Вона стала основою для матеріалістичного розуміння життя, пояснення еволюційного взаємозв'язку організмів, розуміння суті індивідуального розвитку.
Основні положення клітинної теорії зберегли своє значення і нині, хоча більш ніж за 100 років були отримані нові відомості про структуру, життєдіяльність і розвиток клітин.
Сучасна клітинна теорія включає такі положення:
• клітина — елементарна одиниця будови і розвитку всіх живих організмів;
• клітини всіх одно- і багатоклітинних організмів подібні за походженням (гомологічні), будовою, хімічним складом, основними виявами життєдіяльності;
• кожна нова клітина утворюється тільки внаслідок розмноження материнської клітини шляхом поділу;
• у багатоклітинних організмів, які розвиваються з однієї клітини (зиготи, спори тощо), різні типи клітин формуються внаслідок їхньої спеціалізації упродовж індивідуального розвитку особини й утворюють тканини;
• з тканин складаються органи, які тісно пов'язані між собою й підпорядковані нервово-гуморальним та імунним системам регуляції.
Клітинна теорія є одним з найбільших узагальнень природознавства XIX ст. Формування клітинної теорії створило основу для подальших фундаментальних досліджень процесів життєдіяльності, будови та розвитку всіх організмів на Землі.
БУДОВА І ФУНКЦІЇ КЛІТИНИ
Наука, що вивчає будову, хімічний склад, процеси життєдіяльності і розмноження клітин, називається цитологією (гр. куtos — порожнина, 1оgos — наука).
Клітина — це структурна одиниця живих організмів, що є певним чином диференційованою ділянкою цитоплазми, оточеною клітинною мембраною. Функціонально клітина є основною одиницею життєдіяльності організмів.
Клітини існують і як самостійні організми, і входять до складу багатоклітинних організмів. Бактерії, багато видів водоростей {хлорела, хламідомонада), нижчих грибів (цукор, дріжджі) і найпростіші тварини (амеба, евглена зелена, інфузорії тощо) складаються з однієї клітини. Ця клітина виконує всі функції живого організму — живлення, руху, розмноження тощо. Тіло більшості видів рослин і тварин складається з величезної кількості клітин, які спеціалізуються на виконанні окремих функцій. Ці клітини утворюють різні тканини (тканини рослин — див. "Ботаніка", тканини тварин — див. "Анатомія, фізіологія і гігієна людини").
Хоча клітини (рослин, тварин і грибів) мають різну будову і виконують різні функції, вони мають багато спільних морфологічних особливостей (сформоване ядро, подібний набір органел) і подібних функціональних властивостей (біосинтез білків, використання і перетворення енергії, процеси розмноження).
Клітини різняться за розмірами, формою (мал. 2), особливостями організації, функціями. За формою клітини бувають: циліндричні і кубічні (епітеліальні тканини), дископодібні (еритроцити), кулясті (яйцеклітини), видовжені і веретеноподібні (м'язові), зірчасті (нервові) тощо. Серед клітин трапляються і такі, що не мають сталої форми. Це так звані амебоїдні клітини (наприклад, лейкоцити).
Більшість клітин багатоклітинного організму мають розміри від 10 до 100 мкм, а найдрібніші — 2—4 мкм. Великі розміри мають деякі рослинні клітини з великими вакуолями в цитоплазмі: клітини м'якоті кавуна, лимона (їх можна бачити неозброєним оком). Дуже великі розміри (до кількох сантиметрів у діаметрі) мають яйцеклітини птахів і деяких риб. Розміри клітин тваринних організмів не залежать від розмірів їхнього тіла. Наприклад, клітини печінки коня і миші мають майже однакові розміри. Відростки нервових клітин іноді досягають одного і більше метрів. Кількість клітин в організмі зазвичай дуже велика. Лише окремі багатоклітинні організми мають невелику кількість клітин. Наприклад, такі порівняно великі організми, як коловертки, складаються лише з 400 клітин. У хребетних тварин і людини найчисленнішими є клітини крові і головного мозку. Так, у людини кора мозку складається з (14...15) * 109 клітин, а клітин нейроглії у 5—10 разів більше. Кількість еритроцитів у крові дорослої людини становить 23 • 1012. Невеликі розміри і значна кількість клітин створюють у багатоклітинних організмів величезну поверхню, що важливо для забезпечення швидкого обміну речовин.

Мал. 2. Форма клітин організмів людини і тварин:
a — нервова клітина (нейрон); б—г — різні види епітеліальних клітин;
д— клітина сполучної тканини; е — яйцеклітина; є — м'язова клітина
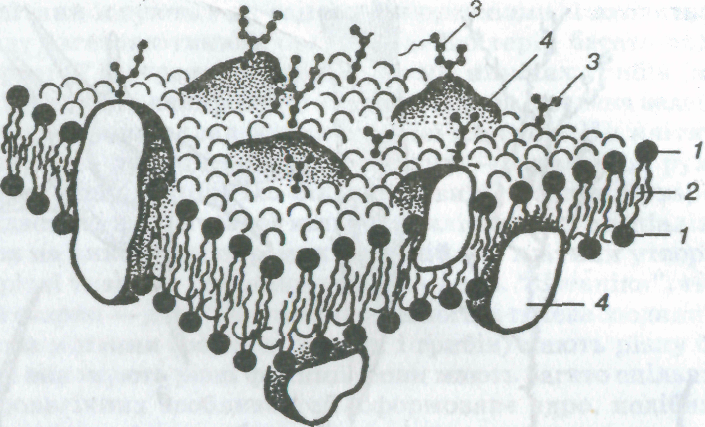
Мал. 3. Мозаїчна модель клітинної мембрани:
1 — гідрофільні головки ліпідів; 2 — гідрофобні частини молекул ліпідів; 3 — молекули вуглеводів; 4 — молекули білків
Зовнішня клітинна мембрана. Зовні клітина вкрита плазматичною мембраною, або зовнішньою клітинною мембраною (мал. З, див. також мал. 1, 5), завтовшки близько 6—10 нм. Це щільна плівка з ліпідів (в основному фосфоліпідів) і білків. Молекули ліпідів розташовані упорядковано — перпендикулярно до поверхні, у два шари так, що ті частини їхньої молекули, які активно взаємодіють з водою (гідрофільні), спрямовані назовні, а частини молекули, які інертні до води (гідрофобні), — всередину мембрани. Молекули білка на поверхні ліпідного каркаса розташовані з обох його боків не суцільним шаром.
Частина молекул білків частково занурена у ліпідний шар, а деякі з них пронизують його наскрізь, утворюючи ділянки, крізь які проникає вода. Ці білки виконують різні функції: одні з них — ферменти, інші — транспортні, які переносять потрібні речовини з навколишнього середовища в цитоплазму і у зворотному напрямку. До молекул ліпідів і білків плазматичної мембрани приєднуються молекули полісахаридів і утворюють тонкий (завтовшки у декілька десятків нанометрів) поверхневий шар клітинної стінки. Його називають глікокаліксом. Він виявлений практично в усіх тваринних клітинах, але ступінь його вираження різний. Глікокалікс забезпечує безпосередній зв'язок клітини із зовнішнім середовищем. У ньому відбувається позаклітинне травлення і розміщені клітинні рецептори.
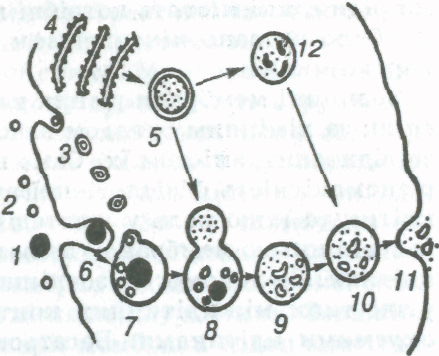
Мал. 4. Лізосомн в процесі фаго- і піноцнтозу:
1,2 — великі і малі часточки в середовищі, яке оточує клітину; захоплення часточок шляхом ліноиитозу (3, 7) і фагоцитозу (6, 7); 4, 5 — утворення лізосо'м (4 — гранулярна ендоплазматична сітка); 8 — виливання ферментів лізосом у вакуолі; 9,10 — розщеплення вмісту вакуолі (лізис); 11 — виділення продуктів розщеплення;12—ділянка цитоплазми після
дії ферментів лізосом.
Однією з основних властивостей біологічної мембрани є її вибіркова проникність (напівпроникність) — деякі речовини проникають крізь неї важко, інші — легко і навіть у бік вищої концентрації. Так, для більшості клітин концентрація іонів Nа+ всередині клітини значно нижча, ніж у навколишньому середовищі. Для іонів К+ характерне протилежне співвідношення: їх концентрація всередині клітини вища, ніж зовні. Тому іони Nа+ завжди намагаються проникнути в клітину, а іони К+ — вийти назовні. Вирівнюванню концентрацій цих іонів перешкоджає особлива система клітинної мембрани, яка виконує роль насоса, що відкачує іони Иа+ з клітини й одночасно накачує іони К+ всередину (так званий натрій-калієвий насос).
У разі активного видалення іонів Na+ з клітини створюються умови для надходження глюкози та амінокислот всередину неї.
У багатьох клітин поглинання речовин відбувається також шляхом фагоцитозу і піноцитозу (мал. 4). Під час фагоцитозу гнучка зовнішня мембрана утворює невеликі заглиблення, куди потрапляє захоплювана тверда часточка. Це заглиблення поступово збільшується, глибшає, часточки, які потрапили в нього, занурюються всередину клітини, й утворюється ендоцитозний пухирець (фагосома, харчова вакуоля). З ним зливається первинна лізосома (що містить неактивовані ферменти), й утворюється вторинна лізосома, або травна вакуоля (гетерофагосома). Злиття супроводжується активацією ферментів і відбувається розщеплення. Продукти розщеплення вбираються і засвоюються цитоплазмою клітини. Рештки виводяться назовні шляхом екзоцитозу. Явище фагоцитозу властиве амебам і деяким іншим найпростішим, а також лейкоцитам (фагоцитам). Аналогічно відбувається і поглинання клітинами рідин, які містять потрібні клітині речовини. Це явище було названо піноцитозом (гр. ріпо — п'ю, куіое — порожнина).
Зовнішні мембрани різних клітин істотно відрізняються як за хімічним складом власних ліпідів і білків, так і за відносним вмістом їх. Саме ці особливості визначають різноманітність фізіологічної активності мембран різних клітин та їхню роль у життєдіяльності клітин і тканин. Із зовнішньою мембраною зв'язана ендоплазматична сітка клітини. За допомогою зовнішніх мембран здійснюються різні типи міжклітинних контактів, тобто зв'язок між окремими клітинами. Багатьом типам клітин властива наявність на їхній поверхні великої кількості виступів, складок, мікро ворсинок. Вони значно збільшують площу поверхні, прискорюють обмін речовин та зміцнюють зв'язки окремих клітин між собою. У рослинних клітинах зовні клітинної мембрани є товсті оболонки, які добре видно в оптичний мікроскоп, що складаються з клітковини — целюлози (див. мал. 1, поз. 11). Ці вторинні клітинні стінки — продукт життєдіяльності органел цитоплазми (зокрема, комплексу Гольджі). Вони створюють міцну опору рослинним тканинам (деревина). Деякі клітини тваринних організмів також мають низку зовнішніх структур, що вкривають мембрану і виконують захисну функцію. Прикладом може бути хітин покривних клітин комах.
Товста целюлозна оболонка не повністю ізолює рослинні клітини одну від одної. У клітинах багатьох рослинних тканин ця оболонка пронизана численними канальцями, крізь які проходять вирости цитоплазми (плазмодесми), що сполучають сусідні клітини. Така структура забезпечує надходження продуктів життєдіяльності з однієї клітини в іншу. Цитоплазма. Внутрішній вміст клітини (протоплазма) ділиться на цитоплазму і ядро. Цитоплазма є основною за об'ємом частиною клітини. За фізичними властивостями це напіврідка маса колоїдної структури, в якій містяться органели клітини мембранної (ендоплазматична сітка, мітохондрії, пластиди, комплекс Гольджі, лізосоми) і не мембранної (рибосоми, центріолі клітинного центру) будови. Агрегатний стан цитоплазми може бути різним: рідким — золь і в'язким — гель. За хімічним складом цитоплазма досить складна.
Під оптичним мікроскопом у цитоплазмі не вдається розгледіти жодних структур, і вона здається однорідною. За допомогою електронного мікроскопа було встановлено, що в цитоплазмі всіх клітин тваринного і рослинного походження є складна система мембран, які часто розміщуються паралельно одна одній. Ця система мембран дістала назву ендоплазматичної сітки. Мембрани завтовшки
6—8 нм мають ліпідно-білкову природу і за структурою
подібні до зовнішньої мембрани клітини. Вони обмежують дуже розгалужену взаємозв'язану систему канальців, щілин і міхурців, яка сполучає різні ділянки клітини. Діаметр порожнини канальців — 25—30 нм. Мембрани ендоплазматичної сітки безпосередньо зв'язані з мембранами комплексу Гольджі. З мембран ендоплазматичної сітки утворюється оболонка ядра після поділу клітини. Зовнішня оболонка ядра є продовженням мембрани ендоплазматичної сітки і навколо ядерний простір сполучений з простором ендоплазматичної сітки.
Ендоплазматичні мембрани бувають двох типів: гладенькі (агранулярні) і шорсткі (гранулярні) (див. мал. 1, поз. 9, 12). Стінки останніх несуть на собі багато рибосом. Ендоплазматична сітка збільшує внутрішню поверхню клітини, що важливо для процесів обміну, бере активну участь у біосинтезі білків, жирів і вуглеводів, транспортує хімічні речовини в різні ділянки клітини.
Крім ендоплазматичної сітки в цитоплазмі є структури не мембранної будови — мікро трубочки. Це тоненькі ниткоподібні структури завтовшки 15—20 нм, до складу яких входять скоротливі білки. Вони утворюють нитки ахромати нового веретена під час мітозу, входять до складу клітинного центру, а також війок і джгутиків клітини. Вважають, що мікро трубочки мають значення для підтримання форми клітини (цитоскелет), а також забезпечують внутрішньоклітинний рух (пересування хромосом) і рух клітини загалом за допомогою джгутиків і війок.
Крім мікро трубочок до складу цитоскелета клітини еукаріотів входять мікрофіламенти та проміжні філаменти. Мікро філаменти складаються в основному з білка актину і трапляються в усіх клітинах еукаріотів. Вони входять до складу м'язових волокон та спеціальних клітинних компонентів (мікро ворсинок), утворюючи пучки в цитоплазмі рухливих клітин тварин. У багатьох рослинних клітинах і клітинах нижчих грибів вони розміщені в шарах рухливої цитоплазми. У клітині вони або є частиною скоротливого апарату, або беруть участь у формуванні жорстких скелетних структур, або утворюють жорстку фібрилярну сітку. Проміжні філаменти побудовані з фібрилярних білків. Це справжня опорна система клітин, що зазнають значних фізичних навантажень (епітеліальні клітини). В аксонах проміжні філаменти (нейрофіламенти) створюють жорстку основу, яка забезпечує гнучкість і цілісність тонких цитоплазматичних відростків нервових клітин. У смугастих м'язах проміжні філаменти входять до складу зет-дисків.
Рибосоми також було виявлено лише за допомогою електронного мікроскопа. Рибосома — органела сферичної форми діаметром 15—20 нм, що складається з молекул рибонуклеїнової кислоти (4 молекули: одна в малій і три у великій субодиницях у еукаріотів, у прокаріотів відповідно 1 і 2) і білка. Кожна рибосома має дві неоднакові субодиниці (субчастинки) — велику і малу (мал. 5), Форма і конфігурація рибосом різних організмів і клітин (прокаріотів і еукаріотів) напрочуд подібні, хоча й різняться в деталях. Утворюються рибосоми в ядерцях у вигляді окремих субодиниць. Субодиниці об'єднуються в рибосому поза ядром. Мала рибосомальна субодиниця має активну ділянку для приєднання матричної і транспортної РНК, а велика — для синтезу поліпептидного ланцюга. Частина рибосом сполучена з мембранами ендоплазматичної сітки (шорстка ЕПС), інша — розміщена в цитоплазмі. Основною функцією рибосом є матричний синтез білків, під час якого рибосоми зв'язують і утримують компоненти бі-
локсинтезуючої системи, виконують каталітичні функції і трансляцію. Особливо багато рибосом у клітинах тканин, які швидко ростуть.
Мітохондрії — орга-нели клітин, які добре видно під оптичним мікроскопом. Вони мають форму паличок, зерен, ниток розміром від 0,5 до 5 мкм. Мітохондрії нервових клітин мають довжину до 20 мкм.
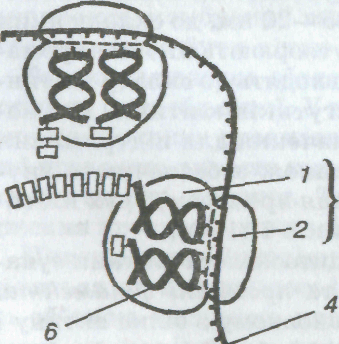
Мал. б. Рибосоми:
1. 2 — відповідно велика і мала субодиниці рибосоми;
З, 4 — інформаційна (матрична) РНК;
5 — поліпептидний ланцюг;
6 — транспортна РНК з амінокислотою
(стрілкою показано напрямок руху іРНК)
Вони є в усіх клітинах, за винятком бактерій і синьо – зелених водоростей. Кількість мітохондрій у клітинах різна — від однієї (у деяких водоростей) до тисячі (печінка). Виявлено, що за певних умов (наприклад, під час голодування) число мітохондрій може зменшуватися. Спостерігаючи під мікроскопом за живими клітинами, можна побачити, що мітохондрії переміщуються в цитоплазмі, можуть утворювати скупчення. У цитоплазмі мітохондрії розмножуються шляхом поділу, можуть також утворюватись внаслідок перетворення мембран ендоплазматичної сітки.
Під електронним мікроскопом було виявлено, що мітохондрії вкриті двома мембранами (зовнішньою і внутрішньою), простір між якими заповнений рідиною. Зовнішня мембрана гладенька. Внутрішня мембрана робить впинання або складки всередину мітохондрії — кристи (лат. сrista — гребінь, виріст), які збільшують внутрішню поверхню. У більшості мітохондрій вони розміщені в поперечному напрямку, деякі з них розгалужені. Кількість крист у мітохондріях різних клітин неоднакова: їх може бути від кількох десятків до кількох сотень, причому особливо багато їх у мітохондріях клітин, які активно функціонують. Внутрішня порожнина мітохондрії заповнена напіврідкою речовиною. В ній містяться рибосоми, РНК і ДНК. На поверхні крист розміщена велика кількість ферментів, які забезпечують перебіг складних біохімічних реакцій. Функція мітохондрій полягає в тому, що в них відбуваються окисно-відновні процеси, внаслідок яких розщеплюються вуглеводи, амінокислоти та інші органічні сполуки. При цьому вивільняється енергія, яка акумулюється шляхом утворення фосфатних зв'язків в аденозинтрифосфорній кислоті — АТФ. Завдяки здатності до нагромадження АТФ мітохондрії є своєрідними акумуляторами енергії клітини. Синтезована в мітохондріях АТФ вільно виходить у цитоплазму й далі прямує до ядра та органел клітини, де в міру потреби розщеплюється і забезпечує їх енергією.
Крім АТФ у мітохондріях синтезуються власні білки, РНК і ДНК.
Пластиди — органели, специфічні для рослинних клітин, а в клітинах тваринних організмів, бактерій, синьо – зелених водоростей і грибів їх немає. У клітинах вищих рослин міститься від 10 до 200 пластид розміром 3—10 мкм, більшість із них має форму двоопуклої лінзи (трапляються у формі паличок, пластинок, лусок і зерен).
Залежно від характеру пігменту розрізняють: хлоропласти (гр. сhioros — зелений) — зеленого кольору, хромопласти — жовтого, оранжевого і червоного кольорів і лейкопласти — безбарвні пластиди. У процесі розвитку рослин пластиди одного типу можуть перетворюватися на пластиди іншого типу. Це явище поширене в природі й особливо помітне під час достигання плодів, коли змінюється їхнє забарвлення. У більшості водоростей пластиди представлені хроматофорами (у клітині він зазвичай один, значних розмірів і має форму сітки, чаші, спіральної стрічки або зірчастої пластинки).
Внутрішня будова пластид досить складна (мал. 6). У хлоропластах є власні рибосоми, ДНК, РНК, включення жиру, зерна крохмалю. Зовні хлоропласти вкриті двома мембранами, всередині заповнені напіврідкою речовиною (строма), в якій містяться особливі, властиві тільки хлоропластам структури — грани. Грани (розміром близько 1 мкм) — пакети круглих плоских мішечків (тилакоїдів), складених подібно до стовпчика монет перпендикулярно до широкої поверхні хлоропласта. Тилакоїди сусідніх гран сполучені між собою мембранними каналами (між гранні ламели) в єдину взаємозв'язану систему. Хлорофіл розміщений у певному порядку на поверхні і в товщі цих мембранних структур хлоропласта. Число гран у хлоропластах різне. Наприклад, у клітинах овочевої культури шпинату кожний хлоропласт містить 40—60 гран.
Подібно до інших органел хлоропласти не закріплені в певних місцях, а здатні змінювати своє положення в клітині шляхом пасивного переміщення разом з течією цитоплазми або шляхом активного орієнтованого переміщення. Активний рух хлоропластів особливо чітко простежується в разі значного посилення однобічного освітлення. При цьому хлоропласти скупчуються біля бічних стінок клітини й орієнтуються до джерела світла ребром. У разі слабкого освітлення хлоропласти орієнтуються більшою площиною до світла й розміщуються вздовж стінки клітини, яка обернена до світла.
За освітлення середньої сили вони займають серединне положення. Цим забезпечуються найсприятливіші умови для процесу фотосинтезу.
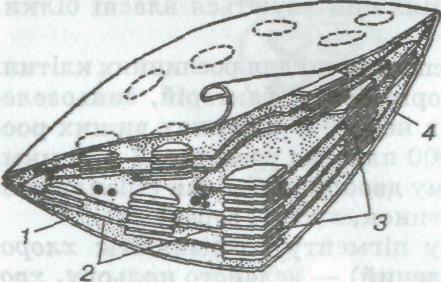
Мал. 6. Схема субмікроскопічної будови хлоропласта:
1 — строма; 2 — ламели; 3 — грани; 4 — оболонка
Складна внутрішня просторова організація структурних елементів хлоропласта сприяє ефективному поглинанню і використанню променистої енергії, а також розмежовує в часі і просторі численні й різноманітні реакції» які в сукупності становлять процес фотосинтезу. Відомо, що залежні від світла (світлові) реакції цього процесу відбуваються лише в тилакоїдах, а біохімічні (темнові) реакції — в стромі хлоропласта.
Молекула хлорофілу дуже подібна до молекули гемоглобіну й відрізняється від останньої в основному тим, що розміщений у центрі молекули гемоглобіну атом заліза замінений у молекулі хлорофілу на атом магнію.
У природі трапляються чотири типи хлорофілу: а, Ь, с, й. Хлорофіли а і Ь містяться у вищих рослинах і зелених водоростях, діатомові водорості містять хлорофіли а і с, червоні — а і d. Краще за інші вивчено хлорофіли а і Ь (вперше їх розділив російський учений М. С. Цвєт на початку XX ст.).
Крім цих хлорофілів існує чотири види бактеріохлорофілів — зелених пігментів пурпурових і зелених бактерій — а, Ь, с, d. Більшість фото синтезуючих бактерій містять бактеріохлорофіл а, деякі — бактеріохлорофіл Ь, зелені бактерії — c i d. Хлорофіл має здатність досить ефективно поглинати променисту енергію і передавати її іншим молекулам. Завдяки цій здатності хлорофіл є єдиною структурою на Землі, яка забезпечує процес фотосинтезу. Пластидам, подібно до мітохондрій, властива автономія в клітині. Вони мають свою власну спадкову інформацію, закодовану у вигляді послідовності нуклеотидів кільцевої молекули ДНК, як і у прокаріотів. Вони мають і власний білоксинтезуючий апарат, за допомогою якого синтезують специфічні білки, що входять до складу їхніх мембран. Як і мітохондрії, пластиди розмножуються поділом.
Поряд із фотосинтезом у хлоропластах відбувається синтез інших речовин, таких як білки, ліпіди, деякі вітаміни.
Завдяки наявності в пластидах ДНК вони відіграють певну роль у передаванні спадкових ознак (цитоплазматична спадковість).
Комплекс Гольджі, або внутрішній сітчастий апарат (див. мал, 1, поз. 7), під оптичним мікроскопом має форму сітки або зігнутих паличкоподібних тілець, розміщених навколо ядра. Трапляється в усіх клітинах тварин і рослин. Особливо добре його видно в нервових клітинах. Під електронним мікроскопом виявлено, що ця органела утворена гладенькими мембранами, які розміщені паралельно одна одній і утворюють систему трубочок з міхурцями різного розміру на кінцях. Розміри дрібних міхурців — 20—30 нм, великих — до 2000 нм.
Основна функція комплексу Гольджі — виведення синтезованих клітиною речовин. Ці речовини транспортуються по канальцях ендоплазматичної сітки і нагромаджуються у міхурцях комплексу Гольджі. Звідси вони або виводяться з клітини назовні, або використовуються в процесі життєдіяльності клітин. У комплексі також відбувається концентрування речовин (наприклад, барвників), які надходять у клітину ззовні і мають бути виведені з неї. Крім того, комплекс Гольджі бере участь у синтезі тих хімічних сполук, з яких будується клітинна мембрана.
Лізосоми (див. мал. 1, поз. 6; мал. 4, поз. 4, 5; гр. lysis — розчинення) — дрібні, видимі тільки під електронним мікроскопом органели діаметром близько 1 мкм. Вони вкриті щільною мембраною і містять до 40 різних ферментів, здатних розщеплювати білки, жири і вуглеводи. Кількість лізосом у клітинах різна. Особливо багато їх (до кількох сотень) у клітинах, здатних до фагоцитозу. Функція лізосом полягає у перетравленні речовин, які потрапили в клітину у процесі фаго- або піноцитозу, а також у руйнуванні окремих органел або клітини у разі ЇЇ відмирання. Це відбувається внаслідок руйнування оболонки лізосом і вивільнення з неї ферментів. Іноді ферменти лізосом беруть участь у руйнуванні міжклітинної речовини, а також цілих органів. Наприклад, під дією ферментів лізосом відбувається розсмоктування хвоста у пуголовків жаби в процесі метаморфозу. Утворення нових лізосом пов'язане з міхурцями комплексу Гольджі.
Клітинний центр (центросома) — органела, видима під оптичним мікроскопом у клітинах тварин, найпростіших та деяких рослин. Розміщена переважно біля ядра. Складається з 1—2, а іноді більше центріолей, оточених щільним шаром цитоплазми — центросферою. Центріоль має форму циліндра завдовжки 0,2—0,3 мкм і діаметром 0,1— 0,15 мкм. Стінка циліндра складається з 9 пар мікротрубочок, утворених фібрилами.
Центросома визначає орієнтацію веретена поділу клітини між центріолями, які розходяться до полюсів клітини, і бере участь у розходженні хромосом до полюсів. З нею пов'язана здатність деяких клітин до активного руху. Це підтверджується тим, що в основі джгутиків або війок рухливих клітин (найпростіші, сперматозоони) містяться
утвори такої самої структури, як і центросоми. У вищих рослин центросом немає. Веретено поділу формується з речовин ядра і цитоплазми клітини (див. "Мітоз").
Органели руху. Клітини можуть переміщуватися за допомогою спеціальних органел, до яких належать війки і джгутики. Війки клітин завжди численні (у найпростіших кількість їх досягає сотень і тисяч), а довжина становить 10—15 мкм. Джгутиків найчастіше буває 1—8, їх довжина становить 20— 50 мкм. Будова війок і джгутиків клітин як рослинних, так і тваринних організмів подібна. За допомогою електронного мікроскопа виявлено, що уздовж них проходять мікротрубоч-ки, дві з яких розміщені в центрі, а навколо них по периферії лежить ще 9 пар мікротрубочок. Уся ця структура вкрита цитоплазматичною мембраною, що є продовженням клітинної мембрани. Рух джгутиків і війок забезпечує не тільки пересування клітин у просторі, а й переміщення різних речовин по поверхні клітин та надходження харчових грудочок у клітину. Біля основи війок і джгутиків розміщені базальні тільця (гомолог центріолі), які також складаються з мікро – трубочок. Вважають, що базальні тільця є центром формування мікро – трубочок джгутиків І війок. Базальні тільця, в свою чергу, нерідко беруть початок від клітинного центру.
Велика кількість одноклітинних організмів і деякі клітини багатоклітинних не мають спеціалізованих органел руху і переміщуються за допомогою псевдоніжок (псевдоподій). Переміщення за допомогою псевдоніжок дістало назву амебоїдного руху. В його основі лежить рух молекул скоротливих білків.
Включення найчастіше трапляються в рослинних клітинах і від органел відрізняються тим, що вони тимчасові — то з'являються, то зникають у процесі життєдіяльності клітини і зазвичай не оточені мембраною. Кількість їх залежить від інтенсивності обміну речовин і стану організму. Під оптичним мікроскопом вони мають форму зерен або крапель різних величини і форми. За хімічним складом розрізняють: вуглеводні, білкові (зерна) та жирові (краплі) включення. У рослинних клітинах вуглеводи найчастіше відкладаються у вигляді зерен крохмалю, а в тваринних — глікогену. Білкові зерна у великій кількості містяться в цитоплазмі яйцеклітин тварин у вигляді жовтка. Багато їх у насінні рослин. Жирові краплі є в клітинах сполучної тканини тварин і в насінні рослин. У рослинних клітинах трапляються кристалічні включення (солі органічних кислот). За певних Умов усі види включень клітина в процесі життєдіяльності може використати, а потім нагромаджувати їх знову.
У цитоплазмі багатьох рослинних і деяких тваринних клітин є особливі органели — вакуолі. У клітинах безхребетних (кишковопорожнинні, війчасті черви, деякі молюски), в одноклітинних організмах та у фагоцитах вищих тварин утворюються травні вакуолі, що містять травні ферменти. У багатьох одноклітинних організмів є ще й пульсівні вакуолі, які, періодично скорочуючись, виводять із клітин продукти розпаду та регулюють осмотичний тиск (осморегуляція). У рослин вакуолі утворюються в процесі росту клітин, Дрібні й численні вакуолі, поступово збільшуючись, зливаються одна з одною й утворюють одну велику вакуолю, що займає майже всю клітину. Речовини клітинного соку (цукри, крохмаль, кислоти тощо), який заповнює вакуолю, сприяють живленню рослин і створюють пружний стан <тургор) у клітинах і тканинах. Особливо великі вакуолі утворюються в старих клітинах.
Ядро. Будова і функції в період інтерфази. Всі клітини тварин (за невеликим винятком — еритроцити) і рослин мають ядро. В більшості клітин є одне ядро, рідше трапляються дво- і багатоядерні клітини. Багатоядерними є клітини деяких видів найпростіших, а також клітини печінки, мозку і м'язів людини. Вони часто виникають внаслідок злиття кількох клітин в одну. Форма ядра здебільшого залежить від форми та розмірів клітини. Зазвичай у кулястих клітинах ядро має округлу форму, у видовжених м'язових клітинах ядро також видовжене. У деяких клітинах ядра можуть мати неправильну форму, наприклад, у лейкоцитів підковоподібні або лапчасті ядра. Форма ядра може змінюатися з віком клітини й залежить від її функціонального стану. Розміри ядра найчастіше коливаються від 2 до 20 мкм. Для кожного типу клітин існує певне ядерно-плазматичне співвідношення, порушення якого призводить до поділу клітини або її загибелі.
Ядро інтерфазної клітини вкрите двома цитоплазматичними мембранами, які відсутні лише в період мітотичного поділу. Зовнішня мембрана ядра часто переходить у мембрани ендоплазматичної сітки і простір між двома ядерними мембранами сполучається з її каналом.
В ядерних мембранах є пори діаметром 80—100 нм. Крізь них відбувається обмін між ядром і цитоплазмою.
Вміст ядра називають ядерним соком (каріоплазмою). У ньому міститься 1—2 ядерця й особлива речовина — хроматин (гр. спгогла — колір, забарвлення). Ця речовина добре фарбується ядерними барвниками. У прокаріотів хроматин складається лише з молекул ДНК, а в еукаріотів — із ДНК,
основних низькомолекулярних білків (гістонів), невеликої кількості кислих білків та іРНК. В інтерфазному ядрі, тобто в період між поділами клітин, хроматин (інтерфазна хромосома) перебуває у вигляді дрібної дифузної зернистості {еухроматин) або тонких ниток і щільних зерен різного розміру (гетерохроматин). Співвідношення еухроматину та гетеро-хроматину залежить від активності процесів у клітині. Чим інтенсивніше відбуваються різноманітні процеси синтезу в клітині, тим більше в них еухроматину, і навпаки. У процесі мітозу в результаті конденсації і скорочення тонких ниток та злиття окремих грудочок хроматину формуються паличкоподібні хромосоми. В період інтерфази в ядрі клітини відбуваються складні процеси біосинтезу ДНК, яка входить до складу хроматину, а також синтезi iРНК (транскрипція — зчитування інформації про структуру білка; див. "Біосинтез білка").
Ядерця мають розміри 0,5—1,0 мкм, містять велику кількість РНК і білка. Вони є місцем синтезу рибосомальної і транспортної РНК, ядерних білків та рибосом. Під час мітозу ядерця зникають, а потім формуються знову в телофазі. Утворення їх пов'язане з функціонуванням певних ділянок хромосом (ядерцевих організаторів), специфічних для кожного виду.
Ядро — це не просто важлива частина клітини, а центр керування її життєвими процесами — обміном речовин, рухом, розмноженням. В ядрі зосереджена основна маса ДНК, яка є носієм спадкової (генетичної) інформації, тобто ядро виконує функцію зберігання інформації про всі ознаки організму, а під час поділу клітини передає її дочірнім клітинам. Позбавлені ядра клітини (наприклад, еритроцити людини) мають порівняно коротку тривалість життя і не здатні до подальшого поділу і відновлення своєї цілісності в разі пошкодження (мал. 7,).
Мал. 7. Одноклітинна зелена водорість ацетабулярія (ілюстрація ролі ядра в регенерації):

a — цілісний організм; 6 — перерізана особина ацетабуля-ріі; в — нижня частина відновлює "шапку" (верхня частина, яка позбавлена ядра, гине).
Таблиця 1. Характерні ознаки клітин прокаріотів та еукаріотів
Показник |
Прокаріоти |
Еукаріоти |
Зовнішня клітинна |
|
|
мембрана |
Є |
Є |
Ядро, вкрите |
|
|
ядерною мембран - |
|
|
ною |
Немає |
Є |
Генетичний апарат |
Одна нитка ДНК, |
Парні хромосоми, |
|
зазвичай замкнена в |
які складаються з |
|
кільце |
комплексу ДНК і |
|
|
білка |
Ендоплазматична |
|
|
сітка |
Немає |
Є |
Рибосоми |
Є (дрібніші) |
Є |
Клітинний центр |
Немає |
Є у більшості тва- |
|
|
рин і деяких рослин |
Мітоховдрії |
Немає |
Є |
Комплекс Гольджі |
Немає |
Є |
Лізосоми |
Немає |
Є |
Джгутики |
Є у деяких видів |
Є у деяких видів, |
|
|
складної будови |
Поділ |
Простий, якому пере- |
Мітоз (непрямий) |
|
дує реплікація ДНК |
|
Прокаріоти та еукаріоти. Будова органел і інтерфазно-го ядра, що описана вище, характерна для клітин усіх тварин і більшості видів рослин та грибів. Ці організми дістали назву ядерних, або еукаріотів. Інша група організмів, менша за чисельністю і, мабуть, давніша за походженням, дістала назву прокаріотів (доядерних). До них належать бактерії та синьо-зелені водорості (їх відносять до царства Дроб'янки), в яких немає справжнього ядра та багатьох органел цитоплазми. Замість ядра у них є нуклеоїд (утвір, подібний до ядра). Він складається із сукупності нуклеїнових кислот і білків (серед них 90 % ДНК і 10 % припадає на білки і РНК), які лежать у цитоплазмі і не відділені від неї мембраною (табл. 1).
ХІМІЧНА ОРГАНІЗАЦІЯ КЛІТИНИ
Вміст хімічних елементів у клітині. Подібність елементарного хімічного складу клітин усіх організмів свідчить про єдність живої природи. Водночас у живих організмах не виявлено жодного хімічного елемента, який би траплявся в тілах неживої природи. Цим підтверджується спільність живої і неживої природи.
Найбільший вміст у клітині чотирьох елементів: кислоти (65—70 %), вуглецю (15—18 %), водню (8—10 %), азоту
(2--3 %).Це органогенні елементи. Разом їх вміст стануть 95—98 % загальної маси живого організму. Вміст у живому організмі таких елементів, як кальцій, калій, фос-фор, сірка, силіцій, натрій, хлор, магній, залізо, становить десяті частки відсотка. Перелічені хімічні елементи належать до макроелементів. Кобальт, цинк, мідь, манган, хром, бром, бор, йод, літій, радій містяться у дуже малих кількостях (менше 0,01 %). їх називають мікроелементами. Важливість того чи іншого хімічного елемента для живих істот визначається не його кількістю. Багато мікроелементів входить до складу ферментів, гормонів та інших життєво важливих сполук, які впливають на процеси розмноження, кровотворення та ін. Наприклад, цинк входить до складу молекули інсуліну; кобальт — до складу ціанкобаламіну (вітаміну В12) тощо.
Усі живі організми значно відрізняються від навколишньої неорганічної природи за кількісним хімічним складом. Наприклад, вуглецю у рослинах міститься близько 18 %, у грунті — менше 1 %, а силіцію, навпаки, у рослинах — 0,15 %, а у грунті — 33 %. Великий вміст вуглецю в складі живих організмів пов'язаний з наявністю в них вуглецевмісних сполук, які називають органічними.
У деяких живих організмах нагромаджуються певні хімічні елементи. Так, у водоростях нагромаджується йод, у жовтці — літій, у болотній рясці — радій тощо.
Вода та інші неорганічні речовини, їхня роль у життєдіяльності клітини. Із неорганічних сполук у клітині найбільше води. Чим вища інтенсивність обміну речовин у тій чи іншій тканині, тим більше вона містить води. В ембріона людини у віці 1,5 місяця вода становить 97,5 %, у вось-мимісячного — 83, у немовляти — 74, у дорослої людини в середньому 66 %. Вміст води в різних органах і тканинах людського організму також різний. Так, мозок дорослої людини містить 86 %, печінка — 70, кістки — 20 % води. З віком вміст води у тканинах зменшується. Вода виконує в клітинах багато функцій: збереження об'єму, забезпечення пружності клітин, розчинення різних хімічних речовин. Крім того, вода — це середовище, в якому відбуваються всі хімічні процеси. Вона безпосередньо бере участь в усіх хімічних реакціях. Так, розщеплення жирів, вуглеводів та інших органічних сполук відбувається в результаті хімічної взаємодії їх з водою. Завдяки високій теплоємності вода захищає цитоплазму від різких коливань температури, сприяє терморегуляції клітин і організму. Частина молекул води (—15 %) у клітинах перебуває у зв'язаному з білковими молекулами стані. Вони ізолюють білкові молекули одну від одної в колоїдних розчинах.
Багато органічних речовин клітини (ліпіди) мають низьку розчинність у воді. Молекули води слабко притягуються до цих речовин, тому вони, становлячи основу клітинної мембрани, обмежують проникнення води з клітини у внутрішнє середовище і навпаки.
Мінеральні солі у великій кількості містяться у клітинах опорних органів — черепашок, хітинових панцирів, кісток. У цитоплазмі інших клітин багато солей перебуває в дисоційованому стані у вигляді катіонів і аніонів — К+, Nа+, Са2+, СI', НСОз, Н?РО4 та ін.
Вміст у клітині катіонів має велике значення для її функціонування. Від концентрації солей залежить надходження води у клітину, бо клітинна мембрана проникна для молекул води і непроникна для багатьох великих молекул та іонів. Якщо в навколишньому середовищі міститься менше іонів, ніж у цитоплазмі клітини, то відбувається надходження води в клітину до вирівнювання концентрації солей (осмос).
Наявність солей у цитоплазмі визначає її буферні властивості — здатність підтримувати стале значення рН (близьке до нейтральної реакції), хоча в процесі обміну речовин безперервно утворюються кислотні й основні продукти.
Будова і біологічні функції органічних речовин, які входять до складу клітини. Різні клітини можуть дуже різнитися за вмістом органічних речовин. У перерахунку на суху масу в клітинах міститься ліпідів — 5—15 %, білків — близько 10—12, вуглеводів — 0,2—2,0, нуклеїнових кислот — 1—2 % маси клітини. Більшість органічних сполук має довгі молекули (полімери), складені з ланцюгів простіших молекул (однорідних або різнорідних мономерів).
Вуглеводи у великій кількості містяться в рослинних клітинах. У деяких плодах, насінні, бульбах вміст їх іноді досягає 90 %. У тваринних клітинах вуглеводів значно менше — до 5 %. Прості вуглеводи називають моносахаридами, складні — полісахаридами. З моносахаридів у організмах трапляються пентози (цикли з 5 атомів вуглецю) і гексози (цикли з 6 атомів вуглецю). Серед пентоз найважливішими є рибоза (складова частина РНК) і дезоксирибоза (складова частина ДНК). Серед гексоз — глюкоза і фруктоза. Вони містяться в багатьох плодах і в
мелУ і зумовлюють солодкий смак їх. Глюкоза є і в крові людини (близько 0,12 %). Цей вуглевод - основний енергетичний матеріал для всіх клітин.
Полісахариди утворюються в процесі полімеризації двох або кількох моносахаридів. Серед дисахаридів найпоширеніші сахароза (складається з молекул глюкози і фруктози) і лактоза, або молочний цукор (складається з молекул глюкози і галактози). З полімерів у природі найчастіше трапляються крохмаль, целюлоза, або клітковина (у рослин), і глікоген (у тварин). Загальна формула їх — (C6H10O5)n, a мономером цих полісахаридів с глюкоза. Наприклад, кожна молекула клітковини утворена ланцюгом із 150—200 молекул глюкози.
Вуглеводи — своєрідне "паливо" для живої клітини: окиснюючись, вони вивільняють хімічну енергію (1г — 17,6 кДж), яка витрачається клітиною на всі процеси життєдіяльності. У рослин вуглеводи виконують і будівельні функції: з них утворюються оболонки клітин. У тварин і грибів у будові клітин бере участь азотовмісний полісахарид — хітин.
Ліпіди — це низькомолекулярні речовини з гідрофобними властивостями. Разом з білками і вуглеводами це основні компоненти всіх видів клітин. У різних органах і тканинах вміст ліпідів неоднаковий. Особливо багато їх у нервовій тканині, серці, печінці, нирках, крові, насінні і плодах деяких рослин.
За хімічною будовою ліпіди досить різноманітні. До складу їхніх молекул входять вищі жирні кислоти, спирти, альдегіди, азотисті основи, амінокислоти, аміноспирти, вуглеводи, фосфорна кислота та ін. Між цими сполуками можуть утворюватися зв'язки: ефірні, складно ефірні, глікозидні, а мідні, фосфоефірні тощо. Класифікація ліпідів дуже складна у зв'язку зі складністю будови молекул цих речовин та їх різноманітністю. Нині всі ліпіди прийнято поділяти на нейтральні (жири) і фосфоліпіди.
Нейтральні ліпіди — це похідні вищих жирних кислот і трьохатомного спирту гліцерину. Як і вуглеводи, жири використовуються як джерело енергії: під час розщеплення одного грама жиру виділяється 38,9 кДж енергії. Підшкірний жир виконує важливу теплоізоляційну функцію, а також сприяє зменшенню впливу ударів та поштовхів. Для тварин, які впадають у сплячку, жири забезпечують організм необхідною енергією, оскільки поживні речовини ззовні в цей час не надходять. Жири становлять запас поживних речовин І в насінні багатьох рослин.
Фосфоліпіди — найбільша частина ліпідів, які входять до складу клітинних мембран. Якщо в середньому на ліпіди припадає 40 % сухої маси мембран, то 80 % з них — на фосфоліпіди. Отже, основні функції мембран (регулювання проникності різних речовин і клітинного вмісту, функціонування іонних насосів, сприйняття, оброблення і передавання всередину клітини інформації з ЇЇ поверхні, імунна відповідь, синтез білків і багато іншого) здійснюються за участю фосфоліпідів (див. мал. 3).
Ліпіди не розчинні у воді і добре розчинні в органічних розчинниках (бензині, ефірі тощо). Самі ліпіди є розчинниками для деяких вітамінів.
Білки, або протеїни, становлять 50—80 % усіх органічних речовин клітини, вони входять до складу міжклітинної рідини, лімфи» плазми крові. Всі види білків мають високу молекулярну масу, яка в окремих випадках досягає 1,5 млн у. о. (умовних одиниць). Всі білки — полімери, мономерами яких є амінокислоти. До складу білків входить близько 20 різних амінокислот. Загальна форму-
™ р — падикал. який може
ла
амінокислот
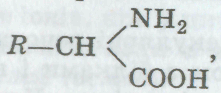 ,
де R
—
радикал, який може
,
де R
—
радикал, який може
СООН
бути циклічною або ланцюговою сполукою. Амінна — МН2 (основна) і карбоксильна — СООН (кислотна) групи визначають амфотерність амінокислот, забезпечують буферність їх розчинів і можливість сполучення їх у довгі ланцюги. Зв'язок між аміногрупою однієї молекули кислоти і карбоксильною групою іншої (виникає в разі відщеплення води) називають пептидним (позначено стрілкою):
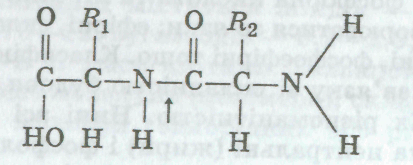 Довгий
ланцюг з амінокислотних залишків,
сполучених пептидними зв'язками (полі
пептид), є первинною
струк* турою білка. У
цьому ланцюзі різні амінокислоти можуть
комбінуватися по-різному. Послідовність
розміщення окремих амінокислот
(первинна
структура білка) визначає
специфічність білків. Заміна, відсутність
або просто перестановка хоча б одного
амінокислотного залишку у поліпептидному
ланцюзі спричинює появу нових білків.
ХІ°'
лі
пептидний ланцюг, утворений кількома
сотнями амінокислотних залишків,
скручується у спіраль, між окремими
витками якої утворюються численні, але
сланкі водневі зв'язки. Така молекула
утворює вторинну
структуру білка. Далі
ця спіраль може скручуватися ще більше
і складатися у клубок,
або
глобулу
(лат.
globus
— куля). У глобулах залишки амінокислот
сполучені слабкими ковалентними
зв'язками. Наприклад, зв'язок S—S
між радикалами двох молекул цистеїну,
які розміщені на великій відстані одна
від одної в поліпептидному ланцюзі. На
цій стадії білок виконує притаманні
йому функції, тобто стає активним.
Довгий
ланцюг з амінокислотних залишків,
сполучених пептидними зв'язками (полі
пептид), є первинною
струк* турою білка. У
цьому ланцюзі різні амінокислоти можуть
комбінуватися по-різному. Послідовність
розміщення окремих амінокислот
(первинна
структура білка) визначає
специфічність білків. Заміна, відсутність
або просто перестановка хоча б одного
амінокислотного залишку у поліпептидному
ланцюзі спричинює появу нових білків.
ХІ°'
лі
пептидний ланцюг, утворений кількома
сотнями амінокислотних залишків,
скручується у спіраль, між окремими
витками якої утворюються численні, але
сланкі водневі зв'язки. Така молекула
утворює вторинну
структуру білка. Далі
ця спіраль може скручуватися ще більше
і складатися у клубок,
або
глобулу
(лат.
globus
— куля). У глобулах залишки амінокислот
сполучені слабкими ковалентними
зв'язками. Наприклад, зв'язок S—S
між радикалами двох молекул цистеїну,
які розміщені на великій відстані одна
від одної в поліпептидному ланцюзі. На
цій стадії білок виконує притаманні
йому функції, тобто стає активним.
Деякі білки утворюють надмолекулярні комплекси, до складу яких входить кілька глобул — це четвертинна структура білка. Наприклад, молекула гемоглобіну складається з чотирьох великих глобул, сполучених між собою також порівняно слабкими зв'язками. Молекули таких білків мають величезну молекулярну масу (кілька мільйонів умовних одиниць).
Під впливом різних фізичних і хімічних факторів можуть відбуватися розкручування білкової молекули і втрата вторинної, третинної та четвертинної структур білка, що призводить до втрати або зміни його властивостей. Порушення специфічної просторової конфігурації білкової молекули має назву денатурації, яка може бути зворотною (у разі збереження первинної структури білка) і незворотною (у разі руйнування первинної структури білка) після припинення дії, яка спричинила зміну його структури.
Значна кількість білків у клітині перебуває у зв'язаному стані з іншими хімічними сполуками. Складові частини таких сполук, як правило, відбиті у їхніх назвах: нуклеопротеїди (нуклеїнова кислота + білок), глюкопротеїд (вуглевод + білок), хромопротеїд (пігмент + білок).
Значення білків дуже велике, бо життя завжди пов'язане з білками. Білки входять до складу всіх органел і мембран клітини, є головним структурним матеріалом.
Дуже важлива рухова функція білків. Комплекси з молекул деяких білків (наприклад, міозину й актину) здатні до скорочення. Завдяки цій властивості білків скорочуються м'язи, рухаються війки і джгутики, переміщуються хромосоми у клітині тощо. Деякі білки виконують в організмі сигнальні функції. З ними пов'язана подразливість клітин і організмів. Ще одна функція білків — захисна, яка забезпечується особливими білками (антитілами), які знешкоджують, нейтралізують побічні (чужорідні) для організму речовини. Нарешті, білки є джерелом енергії. В процесі розщеплення білкової молекули на окремі амінокислоти частина їх може використовуватися для біосинтезу нових молекул білка, а частина розщеплюється повністю, вивільняючи енергію. Внаслідок повного розщеплення 1 г білка вивільняється 17,6 кДж енергії.
Величезне значення мають білки як біокаталізатори, або ферменти.
Ферменти, Молекули простих ферментів складаються лише з амінокислот, а молекули складних можуть функціонувати тільки за наявності в молекулі двох компонентів — білкового (апоферменту) і небілкового (коферменту). Коферментами можуть бути різні органічні речовини, в тому числі і вітаміни, а також метали.
Жодна реакція в клітині не може відбуватися з нормальною швидкістю без участі ферментів як біологічних каталізаторів.
У класифікації ферментів враховують як специфічність їхньої дії на субстрат, так і хімічні реакції, які вони каталізують. Розрізняють ферменти — ліпази (розщеплюють ліпіди), амілази (розщеплюють вуглеводи), пептидази (розщеплюють білки), а також ферменти окисно-відновних реакцій, реакцій гідролізу і синтезу, реакцій перенесення, приєднання або відщеплення певних органічних залишків або груп. Нині складено каталог ферментів, в якому кожному з них присвоєно власний номер і систематичну назву. Наприклад, пепсин за номенклатурою ферментів позначається 3.4.4.1 (пептид пептид о гідролаза), а ліпаза — 3.1.1.3 (гідролаза ефірів гліцерину).
Вибірковість дії ферментів на різні хімічні речовини пов'язана з їхньою будовою. Молекули всіх ферментів мають один або кілька активних центрів, якими вони прикріплюються до тих речовин, на які можуть діяти. Тому дія ферментів завжди специфічна. Наприклад, два травних ферменти — пепсин і трипсин — беруть участь у розщепленні молекул білків до невеликих фрагментів, але кожний з них діє по-різному. Пепсин руйнує зв'язки амінокислоти тирозину, а трипсин — амінокислот аргініну і лізину, причому перший діє на аміногрупи, а другий — на карбоксильні групи амінокислот. Зазвичай ферменти каталізують багато послідовних реакцій, причому речовини, які утворилися за участю першого ферменту, є субстратом для другого тощо. Дія ферментів у клітині завжди узгоджена і відбувається у певній послідовності. Це досягається завдяки тому, що ферменти локалізовані в різних ділянках клітинної мембрани. В органелах клітини ферменти також розміщені послідовно й утворюють упорядковані системи.
Залежно від наявного комплексу ферментів у різних видів організмів і в різних органах обмін речовин відбувається по-різному. Для функціонування кожного ферменту потрібні оптимальні температура і реакція середовища, оскільки одні з них активні в нейтральному середовищі (наприклад, ферменти слини), інші — в кислому (ферменти шлункового соку) або лужному (ферменти підшлункової залози). У разі нагрівання до температури понад 60 °С багато ферментів інактивується (відбувається денатурація білків).
Нуклеїнові кислоти (лат. nucleus — ядро). Ці речовини вперше було виявлено і виділено з ядер клітин. € два види нуклеїнових кислот: дезоксирибонуклеїнова (ДНК) і рибонуклеїнова (РНК). Основна кількість ДНК зосереджена в хромосомах клітини і лише невелика її кількість міститься в мітохондріях і пластидах. РНК міститься в ядерцях, а також у цитоплазмі.
Молекула ДНК — це дуже довгий полінуклеотидний ланцюг, довжина його може досягати десяти міліметрів. Так, вважають, що сумарна довжина молекул ДНК 46 хромосом однієї клітини людини становить 170—180 см. Відповідно дуже велика і молекулярна маса ДНК (сотні мільйонів умовних одиниць).
Кожна молекула ДНК складається з двох сполучених між собою ланцюгів нуклеотидів. До складу кожного нук-леотиду входять азотиста основа, дезоксирибоза і фосфорна кислота. Всього в ДНК є чотири види азотистих основ: аденін (А), гуанін (Г), тимін (Т) і цитозин (Ц) (мал. 8).
Нуклеотиди різняться лише азотистими основами. Назва нуклеотидів також пов'язана з назвою нуклеозидів (сполук азотистих основ з пентозою) цих основ. Наприклад, нуклеотид, який містить аденін, утворюється приєднанням залишку фосфорної кислоти до аденозинового нуклеозиду (аденін + рибоза або дезоксирибоза), називається аденіловим. Відповідно утворюються нуклеотиди гуанозинового, Уридинового, цитидинового і тимідинового нуклеозидів.
Два ланцюги, які складаються з десятків і сотень нуклеотидів, скручуються між собою й утворюють подвійну спіраль. Діаметр цієї спіралі становить 2 * 10-9 м (2 нм), а відстань між сусідніми нуклеотидами — 3,4 • 10-10 м (034 нм; 1 нм = 10-9 м). Розмір витка спіралі (крок спіралі) становить 3,4 • 10-9 м (3,4 нм), у ньому розміщується 10 пар нуклеотидів.
Азотисті основи одного ланцюга сполучені з основами другого ланцюга за допомогою водневих зв'язків у такому порядку: аденін одного ланцюга сполучений з тимі-ном другого ланцюга; гуанін сполучений так само з цитозином
Сполучення інших типів у нормі ніколи не виникають. Це пояснюється тим, що між такими парами нуклеотидів, як аденін" — тимін і гуанін — цитозин, існує особливий тип зв'язку, який дістав назву комплементарного {принцип комплементарності; лат. сотріетепіиш — доповнення).
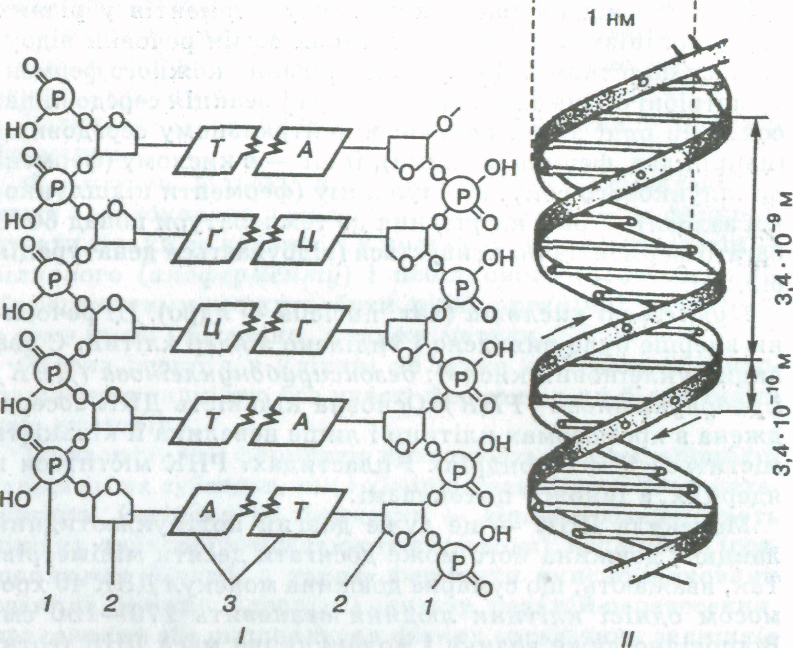
Мал. 8. Схема будови молекули ДНК (/) та її спіральної структури (//):
1— залишок фосфорної кислоти; 2 — дезоксирибоза, 3 — азотисті
. Сполучення інших типів у нормі ніколи не виникають. Це пояснюється тим, що між такими парами нуклео-тидів, як аденін" — тимін і гуанін — цитозин, існує особливий тип зв'язку, який дістав назву комплементарного {принцип комплементарності; лат. сотріетепіиш — доповнення). При цьому між А і Т утворюється два водневих зв'язки, а між Г і Ц — три. Знаючи послідовність сполучення нуклеотидів одного ланцюга молекули ДНК, можна встановити порядок розміщення нуклеотидів другого. Наприклад, якщо в одному ланцюзі послідовність нуклеотидів буде А—А—А—Ц—Т—Т—Г—Г—Г, то на відповідній ділянці другого ланцюга послідовність обов'язково буде Т—Т—Т^-Г—А—А—Ц—Ц—Ц. Приклад розв'язування задач до цього розділу див. у додатку — задача 2.
Подвійна спіраль молекули ДНК здатна розкручуватися, при цьому водневі зв'язки розриваються й окремі ланцюги ДНК відходять один від одного. Іноді таке роз'єднання ланцюгів відбувається не по всій довжині молекули, а лише на певній ділянці. Ланцюги ДНК можуть знову спаралізуватися, відновлюючи свою попередню структуру.
Редуплікація (лат. duplicatio — подвоєння), реплікація (англ. replica — відбиток) ДНК — процес самовідтворення макромолекул нуклеїнових кислот, який забезпечує точне копіювання генетичної інформації і передавання її з покоління в покоління. Самоподвоєння молекули ДНК відбувається в період інтерфази перед поділом клітин. При цьому молекула ДНК розкручується і з одного кінця спіраль розділяється на окремі ланцюги. Біля кожного з них із вільних нуклеотидів, які є в ядрі клітини, розпочинається синтез другого ланцюга ДНК. Цей синтез відбувається за принципом комплементарності. В результаті замість однієї молекули ДНК утворюються дві молекули такого самого нуклеотидного складу, як і початкова. Один ланцюг у кожній новоутвореній молекулі ДНК походить від початкової молекули (материнський ланцюг), а другий синтезується заново (дочірній). Як процес розділення молекули ДНК на два ланцюги, так і процес синтезу нових ланцюгів здійснюється за рахунок дії низки ферментів (ДНК-полімерази, ДНК-лІгази).
Молекула РНК має простішу будову. Вона складається з одного полінуклеотидного ланцюга, який теж містить багато нуклеотидів чотирьох видів — гуаніловий, цитиди-ловий, аденіловий та уридиловий (у молекулі РНК замість основи тиміну міститься азотиста основа урацил, яка за структурою близька до тиміну). До нуклеотидів РНК входить не дезоксирибоза, а рибоза. Молекули РНК значно коротші, ніж молекули ДНК, і мають набагато меншу молекулярну масу, яка рідко перевищує 100 тис. у. о. Існує кілька видів РНК, що різняться за структурою і функціями. Так, рибосомальні РНК (рРНК) містяться в рибосомах, транспортні РНК (тРНК), найбільші за розміром, беруть участь у транспортуванні амінокислот до місця синтезу білків, інформаційні, або матричні РНК (ІРНК, або мРНК), синтезуються на ділянці одного з ланцюгів ДНК і передають інформацію про структуру білка з ядра клітини до рибосом.
Аденозинфосфорні кислоти. Нуклеотиди в клітині трапляються не лише як структурні елементи нуклеїнових кислот, а й як речовини, що функціонують самостійно. Найважливішими в життєдіяльності клітини є аденілові нуклеотиди — моно-, ди- і трифосфорні ефіри аденозину,які містять аденін, рибозу та один (аденозинмонофосфор-на, АМФ), два (аденозиндифосфорна, АДФ) або три (адено-зинтрифосфорна, АТФ) залишки фосфорної кислоти. Ці сполуки є у всіх живих організмах і відіграють величезну роль в енергетичному і пластичному обміні. Приєднання фосфатних залишків до АМФ супроводжується акумулюванням енергії, а гідролітичне відщеплення їх — виділенням енергії. Вивільнена енергія використовується в процесі життєдіяльності клітин. АДФ і АМФ утворюються внаслідок дефосфорування АТФ у процесі фотосинтезу, дихання або гліколізу.
АТФ — це універсальна макроергічна сполука, в якій два із трьох залишків фосфорної кислоти — високоенергетичні (макроергічні). Один із них або обидва легко відщеплюються під впливом ферментів, що супроводжується вивільненням енергії, яка використовується для забезпечення перебігу різноманітних процесів у клітині. Відщеплення 1 моль кислотних залишків фосфорної кислоти супроводжується виділенням майже 40 кДж енергії':

Контрольні запитання і заслання
1. Назвіть основні положення клітинної теорії. Хто їх сформулював?
2. Дайте визначення поняття "клітина". Чому клітина є основною структурною і функціональною одиницею живого?
3. Схарактеризуйте форму, розміри та кількість клітин у багатоклітинних організмах.
4. Перелічіть структурні компоненти рослинних і тваринних клітин, Яка відмінність у будові цих клітин?
5. Як побудована зовнішня клітинна мембрана?
6. Схарактеризуйте будову інтерфазного ядра клітини. Для чого потрібні ядерця?
7. Яке значення ядра в клітині?
8. Що таке хроматин? Яку функцію він виковує?
9. Яку будову мають органели клітини?
10. Коротко схарактеризуйте функції кожного виду органе.7].
11. Які є види клітинних включень І чим вони відрізняються від органел?
12. Перелічіть особливості будови клітин прокаріотів і еука-ріотів.
13. Як надходять у клітину хімічні речовини?
14. Перелічіть основні хімічні макро- і мікроелементи, що входять до складу живих організмів.
15 Яка роль води та інших неорганічних сполук клітини?
16. Назвіть основні органічні сполуки, які входять до складу цитоплазми. Що таке біополімери?
17. Схарактеризуйте будову і біологічне значення вуглеводів і жирів клітини.
18. Чим характеризуються амінокислоти і як вони сполучаються в білковій молекулі?
19. Схарактеризуйте будову білків у клітині. Як утворюються первинна, вторинна І третинна структури білка?
20. Яке значення білків у клітині і в організмі?
21. Як побудовані ферменти та які функції вони виконують?
22. Порівняйте будову молекул ДНК і РНК.
23. Що таке комплементарність у розміщенні нуклеотидів молекули ДНК і яке значення має це явище?
24. Як відбувається подвоєння молекул ДНК?
25. Які види молекул РНК є в клітині та які їхні функції?
26. Що таке АТФ і яке її біологічне значення?
Обмін речовин і перетворення енергії в клітині
Обмін речовин — загальна властивість, характерна для всіх живих організмів.
Загальнобіологічна суть обміну речовин як специфічної властивості живої матерії полягає в тому, що всі живі організми вилучають з навколишнього середовища різні органічні і неорганічні сполуки та хімічні елементи, використовують їх у своїй життєдіяльності і виділяють у зовнішнє середовище кінцеві продукти обміну у вигляді простіших органічних і неорганічних сполук. Обмін речовин можна схарактеризувати як комплекс біохімічних і фізіологічних процесів, які забезпечують життєдіяльність організмів у тісному взаємозв'язку з навколишнім середовищем. Комплекс фізіологічних процесів, що вивчається на рівні цілісного вищого організму, охоплює акти дихання, живлення, травлення, всмоктування, а також виділення продуктів обміну органами і системами (шкіра, легені, видільна система, травний апарат).
Пластичний і енергетичний обмін. Біохімічні процеси, які вивчаються на рівні тканин і клітин, охоплюють хімічні перетворення і видозміни структур білків, жирів і вуглеводів, що надходять в організм у вигляді їжі. Всі ці процеси відбуваються за участю великої кількості ферментів, які забезпечують певну послідовність обмінних реакцій у часі, місце і швидкість перебігу їх. Одночасно з процесом розкладання складних органічних сполук на простіші у клітині відбуваються також процеси синтезу складних органічних сполук (біологічний синтез, або біосинтез). Цим терміном позначають усі біохімічні процеси, які відбуваються в живих організмах і супроводжуються утворенням із простих, низькомолекулярних речовин складних високомолекулярних сполук (нуклеїнових кислот, білків, полісахаридів). Основні хімічні сполуки (амінокислоти, нуклеотиди тощо) синтезуються в клітині із глюкози та аміаку в результаті перебігу кількох сотень послідовних хімічних реакцій. Кожний етап у цій послідовності реакцій здійснюється специфічним ферментом. Отже, глюкоза — це джерело енергії в клітині й основна хімічна сполука для синтезу найважливіших органічних речовин. Процеси розщеплення і синтезу в клітині узгоджені так, що близько половини вуглецевих атомів входить до складу різних хімічних сполук, а решта окиснюеться до вуглекислого газу.
Сукупність усіх реакцій біосинтезу прийнято називати асиміляцією (анаболізмом) (лат. assimilatio — уподібнення), або пластичним обміном. Усі реакції пластичного обміну відбуваються з поглинанням енергії (ендотермічні). Протилежний процес — розщеплення й окиснення органічних сполук у клітині — має назву дисиміляції (катаболізму) (лат. dissimilatio — робити несхожим), або енергетичного обміну. Всі реакції енергетичного обміну відбуваються з виділенням енергії (екзотермічні).
Реакції, які відбуваються під час асиміляції і дисиміляції, хоча й протилежні, однак у живих організмах тісно взаємозв'язані і невіддільні одні від одних. Вони становлять дві сторони єдиного процесу обміну речовин.
Обмін речовин І енергії в клітині (сукупність пластичного та енергетичного обмінів) — це основна умова підтримання життя клітини, основа її функціонування й розвитку.
Етапи енергетичного обміну. Для життєдіяльності клітини, її функціонування, росту, розмноження, синтезу органічних сполук необхідна енергія. Основним джерелом отримання енергії клітиною є глюкоза. Якщо у клітину потрапляє не глюкоза, а якийсь інший вуглевод, то він перетворюється на глюкозу або на одну із проміжних сполук, які утворюються в процесі розщеплення глюкози, а далі ці речовини розщеплюються подібно до глюкози. У цьому процесі високомолекулярні органічні сполуки перетворюються на прості органічні і неорганічні. Процес енергетичного обміну дуже складний. Схематично він може бути зведений до двох етапів.
Як відомо, в процесі травлення складні органічні сполу-розщеплюються в травному каналі на простіші:— на гліцерин і жирні кислоти, полісахариди — на моносахариди, білки — на амінокислоти. Всмоктуючись, вони надходять до внутрішнього середовища організму. Цей процес іноді розглядають як підготовчий етап енергетичного обміну речовин в організмі. Він супроводжується виділенням дорівняно невеликої кількості енергії, яка розсіюється у вигляді теплоти.
На першому етапі енергетичного обміну речовини, які утворилися під час підготовчого етапу, включаються в подальший процес розщеплення без участі кисню. Це складний, багатоступінчастий процес, який відбувається на внутрішньоклітинних мембранах, де є відповідні ферменти. Речовини переміщуються по цих ферментах, як по конвеєру. Розглянемо це на прикладі розщеплення глюкози, яке має спеціальну назву — гліколіз. У процесі гліколізу кисень участі не бере, тому його називають без кисневим розщепленням. Реакція гліколізу в клітині відбувається за участю фосфорної кислоти та АДФ. Сумарне рівняння має такий вигляд:
СвНі206 + 2Н3РО4 + 2АДФ -» 2С8НвО3 +
+ 2АТФ + 2Н2О + 200 кДж.
80 кДж (акумулюється 120 кДж (розсіюється
в АТФ) у вигляді теплоти)
Але видно з рівняння, в процесі гліколізу в АТФ акумулюється близько 40 % енергії (80 кДж із 200). Процес гліколізу відбувається у клітинах тваринних організмів, молочнокислих бактерій, а також у деяких грибів. У клітинах більшості рослин без кисневе розщеплення речовин відбувається шляхом спиртового бродіння. Багато стадій цього процесу аналогічні гліколізу, але кінцевими продуктами його замість молочної кислоти є вуглекислий газ і етиловий спирт (С2Н5ОН).
Другий етап енергетичного обміну — повне, або кисневе, розщеплення. Основною умовою етапу є надходження в клітину достатньої кількості кисню. Як і гліколіз, кисневе розщеплення — це низка послідовних реакцій, кожну з яких каналізує певний фермент. Усі ці процеси відбуваються на мембранах мітохондрій. Проміжні реакції розщеплення молочної кислоти до кінцевих продуктів (СО2 і Н2О) відбуваються з виділенням енергії. Поступовість кисневого розщеплення і виділення енергії надзвичайно важлива для акумулювання енергії в АТФ. У цьому процесі також беруть участь фосфорна кислота та АДФ.
Сумарне рівняння кисневого розщеплення можна запи-ги так:
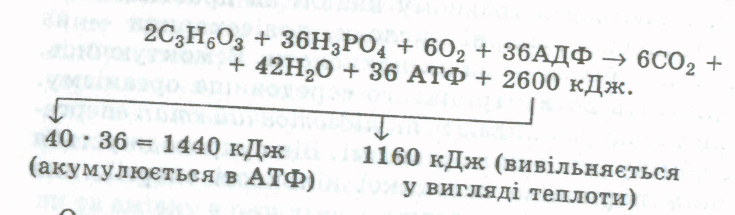
Отже, на другому етапі енергетичного обміну в АТФ акумулюється близько 55 % виділеної енергії (1440 кДж із 2600).
Порівняння без кисневого і кисневого етапів дає змогу побачити, що останній значно ефективніший. Кількість виділеної й акумульованої енергії значно більша, ніж у разі без кисневого розщеплення. Обидва ці процеси розщеплення зумовлюють акумуляцію 1520 кДж енергії, яка зосереджується в 38 молекулах АТФ.
Приклад розв'язування задач із цього розділу див. у додатку.
Синтезована в мітохондріях (або хлоропластах) АТФ по каналах ендоплазматичної сітки надходить у рибосоми та інші ділянки клітини. Там АТФ перетворюється на АДФ, віддаючи акумульовану енергію на синтез білків, ліпідів, вуглеводів, ДНК, скорочення м'язів, поділ клітин та інші потреби організму. Цим самим АТФ здійснює функцію транспорту енергії в організмі. Цей процес можна проілюструвати такою схемою:
 Автотрофні
та гетеротрофні організми. В процесі
історичного розвитку (філогенезу) у
кожного виду живих організмів виробився
власний, особливий тип обміну речовин.
За характером живлення і використання
енергії в процесі обміну речовин усі
організми поділяють на дві групи. 42
Автотрофні
та гетеротрофні організми. В процесі
історичного розвитку (філогенезу) у
кожного виду живих організмів виробився
власний, особливий тип обміну речовин.
За характером живлення і використання
енергії в процесі обміну речовин усі
організми поділяють на дві групи. 42
Організми першої групи — автотрофи (гр. autos
caм , trophe — живити) — здатні синтезувати органічні речовини з неорганічного вуглецю (СО2), використовуючи для нього енергію Сонця (фото синтезуючі організми) або енергію екзотермічних реакцій окиснення неорганічних речовин (хемосинтезуючі). До фото синтезуючих автотрофів належать усі зелені рослини, тобто організми, які містять хлорофіл. До хемосинтезуючих автотрофів належить невелика кількість видів бактерій (наприклад, нітрифікуючи бактерії, залізо і сіркобактерії; див, "Бактерії").
Друга група — гетеротрофи — для синтезу власних органічних сполук використовують вуглець у формі інших органічних сполук, які для них є як джерелом енергії, так і будівельним матеріалом. До таких організмів належать усі тварини, гриби, більшість бактерій та деякі рослини. Якщо гетеротрофи отримують готові органічні речовини від живих організмів, то їх вважають хижаками або паразитами, а якщо вони використовують речовини відмерлих організмів, — то сапрофітами. До гетеротрофних відносять і організми, які живляться за рахунок інших, але не паразитують на них, а перебувають з ними у взаємовигідних відносинах (мутуалізм).
Фотосинтез, його світлова і темнова стадії. Фотосинтезом називають процес синтезу органічних сполук з неорганічних (СО2 та Н2О), який відбувається з використанням променистої енергії Сонця за участю хлорофілу.
Цей складний і багатоступінчастий процес (мал. 9) розпочинається з поглинання квантів світла молекулою хлорофілу. Зелений колір його зумовлений поглинанням переважно червоних і фіолетових променів сонячного спектра. З моменту поглинання сонячного світла хлорофілом розпочинається світлова стадія фотосинтезу.
АТФ
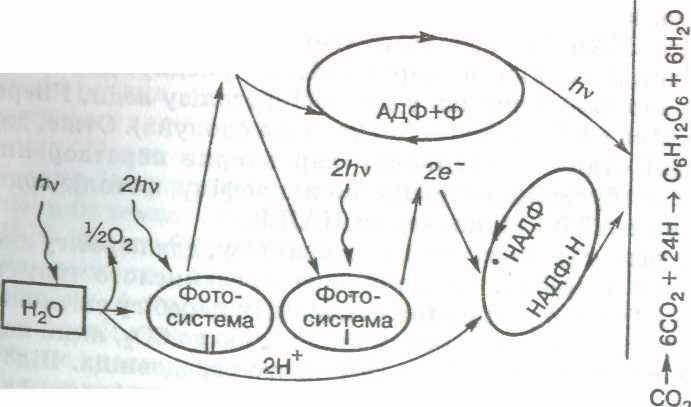
Світлова стадія Темнова стадія
Мал. 9. Схема фотосинтезу
Під впливом фотонів світла відбувається збудження молекули хлорофілу, причому рівні збудження можуть бути різними. Суть цього процесу полягає в тому, що електрони в молекулі хлорофілу переходять на вищий енергетичний рівень, нагромаджуючи потенціальну енергію. Частина з них відразу повертається на попередній рівень, а енергія, яка виділяється при цьому, випромінюється у вигляді теплоти. Значна частина електронів з високим рівнем енергії передає її іншим хімічним сполукам для виконання фотохімічної роботи, яка здійснюється в кількох основних напрямках.
1. Перетворення енергії електронів на енергію АТФ: АДФ .+ Ф + Енергія —> АТФ. Оскільки приєднання залишків фосфорної кислоти відбувається за рахунок енергії світла, цей процес називається фотофосфорилюваннлм.
2. Перебіг процесу фотолізу води: Н2О —> Н+ + ОН~. У результаті іони Н+, приєднуючи електрони з високим енергетичним рівнем, перетворюються на атомарний водень, який використовується в наступних реакціях фотосинтезу, а гідроксильні іони, взаємодіючи між собою, утворюють молекулярний кисень, воду і вільні електрони:
4ОН--* 2Н2О + 02 + 4е'.
3. Передача енергії електронами з високим енергетичним рівнем через низку проміжних речовин для відновлення універсального біологічного переносника (акцептора) водню НАДФ (нікотинамідаденіндинуклеотидфосфат). Внаслідок поглинання енергії НАДФ приєднує два атоми водню, що вивільнились у процесі фотолізу води, і перетворюється на НАДФ • Н2 (відновлена сполука). Отже, для світлової стадії фотосинтезу характерне перетворення енергії — збудження електронів хлорофілу, фотоліз води, утворення АТФ і відновлення НАДФ.
Далі настає темнова стадія фотосинтезу, для перебігу якої світло не потрібне. За наявності вуглекислого газу та енергії АТФ, що утворилися внаслідок перебігу світлових реакцій, відбувається приєднання водню до С02, який надходить у хлоропласти із зовнішнього середовища. Відбуваються послідовні реакції за участю специфічних ферментів, внаслідок чого утворюються різні сполуки, серед яких перше місце посідають вуглеводи. 44
Процес фотосинтезу можна подати таким сумарним рівнянням:
+ Енергія
6СО2 + 6Н2О-------------С6Н,2Об + 6О2.
Фотосинтез має велике значення для існування біосферний Зелені рослини завдяки фотосинтезу щорічно вносять до складу органічних речовин близько 170 млрд т вуглецю здатні поновити увесь кисень атмосфери приблизно за 2 тис. років і увесь вуглекислий газ — за 300 років. Проте в процесі фотосинтезу використовується лише 1 % усієї сонячної енергії, яка потрапляє на рослини.
Вагомий внесок у вивчення ролі світла і хлорофілу в процесі фотосинтезу зробив видатний російський вчений К. А. Тимірязєв. За його словами, зелені рослини відіграють космічну роль завдяки тому, що вони здатні засвоювати сонячну енергію. Ця енергія, акумульована в органічних речовинах, використовується всіма живими організмами нашої планети.
Шляхи підвищення продуктивності фотосинтезу. Складні біохімічні процеси, які відбуваються під час світлової і темнової стадії фотосинтезу, зумовлюють і складний характер залежності цієї функції від умов життя рослини. На інтенсивність процесу фотосинтезу впливають як комплекс зовнішніх факторів — освітленість, температура середовища, вміст вуглекислого газу, вологість тощо, так і біологічні особливості рослин, специфіка їхньої реакції на зовнішні впливи. Ось чому процес фотосинтезу слід розглядати як результат взаємодії всього комплексу внутрішніх і зовнішніх чинників у життєдіяльності рослин. Щодо освітленості, температури, вологості потреби різних видів рослин дуже відрізняються — є світлолюбні і тіневитривалі види, теплолюбні, холодостійкі, посухостійкі види тощо. Проте можна зазначити, що для більшості видів рослин інтенсивність фотосинтезу посилюється з підвищенням температури і досягає максимуму за температури л5 С, вмісту СО2 близько 1 % і насичення водою. Подальше зростання цих показників може дослаблювати інтенсивність фотосинтезу. Підвищення Інтенсивності сонячного освітлення від 1 до 30 % (від максимального) спричинює значне посилення інтенсивності фотосинтезу в усіх вищих рослин, а подальше підвищення інтенсивності освітлення посилює фотосинтез лише у світлолюбних рослин. Фотосинтез — це основний процес утворення органічних речовин, що в поєднанні з асиміляцією мінеральних
солей із ґрунту створює біомасу рослин. Органічні речовини, що утворюються в процесі фотосинтезу, становлять близько 95 % сухої маси рослини. Тому керування процесом фотосинтезу, підвищення його продуктивності — один із ефективних методів впливу на продуктивність рослин, а для сільськогосподарських культур — це важливий засіб підвищення врожаю. Розроблено комплекс агротехнічних заходів, які дають змогу впливати на процес фотосинтезу. До них належить забезпечення потреб рослини водою і мінеральними солями, у тому числі мікроелементами (міддю, цинком тощо), від яких залежить продуктивність роботи всього фото синтезуючого апарату рослин. Дуже ефективним методом є підвищення вмісту СО2 шляхом поливання рослин водою, яка насичена вуглекислим газом. Важливим є також правильне розміщення рослин та густоти посіву їх. Цей метод дає змогу запобігти само затіненню рослин і використати максимальну площу їхніх листків. Велику роль в ефективності використання сільськогосподарськими культурами сонячної енергії відіграє селекція — створення посухостійких сортів, які мають високі інтенсивності фотосинтезу і ростових процесів. Код ДНК. Поняття про ген. Найважливішим досягненням біології XX ст. стало з'ясування генетичного коду — встановлення відповідності між послідовністю нуклеотидів молекули ДНК та амінокислотами молекули білка. Нині генетичний код з'ясовано повністю. Кожна амінокислота кодується трьома (розміщеними поряд) нуклеотидамя молекули ДНК або відповідними (комплементарними) нуклеотидами інформаційної РНК. Ці нуклеотиди складають триплети (трійки, кодони). Чотири різні нуклеотиди молекули ДНК — А, Ц, Т, Г (або А, Ц, У, Г молекули РНК) можуть утворювати 64 різних триплети (з урахуванням послідовності розміщення). Всі ці триплети (за винятком трьох; УАА, УАГ і УГА) відповідають 20 амінокислотам, які входять до складу білків. Деякі амінокислоти, наприклад триптофан (УГГ), метіонін (АУГ), кодуються лише одним триплетом, інші — двома (фенілаланін — УУУ, УУЦ; цистеїн — УГУ, УГЦ), трьома (ізолейцин — АУУ, АУЦ, АУА), чотирма (гліцин — ГГУ, ГГЦ; ГГА, ГГГ; пролін — ЦЦУ, ЦЦЦ, ЦЦА, ЦЦГ) і шістьма (се-рин — УЦУ, УЦЦ, УЦА, УЦГ, АГУ, АГЦ) триплетами. Як видно з наведених прикладів, у разі кодування амінокислот кількома триплетами ці триплети відрізняються лише третьою літерою. У разі шести триплетів чотири з них відрізняються лише третьою літерою, а два відрізняються
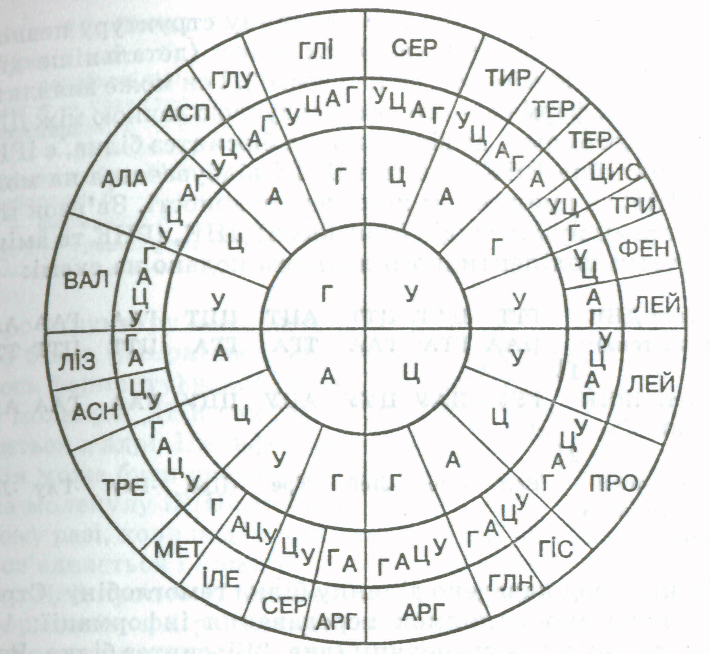
Мал. 10. Генетичний код (РНК):
три азотисті основи в центральному, другому і третьому колах кодують одну амінокислоту, яка скорочено записана у зовнішньому колі
(від цих чотирьох) повністю, але між собою вони відрізняються теж лише третьою літерою (диз. код амінокислоти серину). Триплети УАА, УАГ і УГА (на мал. 10 вони позначені абревіатурою "ТЕР") виконують функцію "розділових знаків" і не несуть генетичної інформації. Вони відділяють інформативні ділянки одну від одної, є стоп-кодо-нами. Саме на них припиняється синтез одного поліпептид-ного ланцюга. Очевидно, стоп-кодон — це кінцева точка функціональної одиниці ДНК-цистрона. Починається синтез наступного ланцюга триплетом АУГ або ГУГ.
Генетичний код характеризується виродженістю (кількість амінокислот менша від кількості триплетів). Він однозначний (кожен триплет кодує Лише одну певну амінокислоту), універсальний (єдиний для всіх організмів) і не перекривається.
Так, на довгому ланцюзі молекули ДНК закодована інформація про структуру різноманітних білків. Подібний код є універсальним, оскільки він однаково функціонує в усіх організмів — від вірусів до людини. Ділянку ДНК,
яка містить інформацію про первинну структуру певного білка, називають структурним геном (детальніше див. "Основні закономірності спадковості"). Ген може виявлятися в кількох формах — алелях. Сполучною ланкою між ДНК ядра і рибосомами, де відбувається біосинтез білка, є ІРНК (інформаційна РНК). Синтез ІРНК відбувається на молекулі ДНК за принципом комплементарності. Зв'язок між триплетами нуклеотидів (кодонами) ДНК, ІРНК та амінокислотами поліпептидного ланцюга подано на схемі:
Триплети ДНК ГТТ ДАТ ЦТТ АЦТ ЦЦТ ГАА ГАА ААА
(частина гена) ЦАА ГТА ГАА ТГА ГГА ЦТТ ЦТТ ТТТ
Триплети ІРНК ГУУ ЦАУ ЦУУ АЦУ ЦЦУ ГАА ГАА ААА
(кодони)
Амінокислоти Вал -Гіс -Лей -Тре -Про -Глу -Глу -Ліз
поліпептидного
ланцюга
Як приклад наведено ділянку білка гемоглобіну- Стрілками показано напрямок передавання інформації: 1 — транскрипції; 2 — трансляції (див, "Біосинтез білка. Роль нуклеїнових кислот"). Як видно зі схеми, транскрипція (синтез іРНК) відбувається лише з одного ланцюга ДНК.
Приклад розв'язування задач такого типу див. у додатку (задача 3).
Біосинтез білка. Роль нуклеїнових кислот. Універсальним для всіх живих організмів видом пластичного обміну є процес біосинтезу білка. Цей процес інтенсивно відбувається в період росту і розвитку організму (збільшення маси організму), а також у тих клітинах, які синтезують ферменти, гормони та інші білкові речовини. У всіх інших клітинах біосинтез іде менш інтенсивно, але триває постійно, бо в клітинах регулярно відбувається розщеплення білків і їх потрібно поновлювати. Найважливішу роль у процесі біосинтезу білка відіграють нуклеїнові кислоти — РНК і ДНК. Сама ДНК безпосередньої участі в синтезі білка не бере, оскільки вона перебуває в ядрі, а основним місцем синтезу білка є рибосоми на ендоплазматичній сітці цитоплазми. Принцип комплементарності, який визначає будову подвійного ланцюга ДНК, лежить в основі і матричного біосинтезу білка. В цьому процесі можна виділити чотири етапи (див. мал. 5, 11).
І етап — транскрипція — передавання інформації про структуру білка з молекули ДНК на іРНК.
Мал. 11- Схема біосинтезу білка в клітині:
a
—
реплікація;
б
— транскрипція;
в
— трансляція;
1
— ДНК-полімераза;
2
—
РНК-полімера-за;
3
—
рибосомальна РНК; 4
— інформаційна
РНК (ІРНК); 5 -рибосоми; 6
— білкові
ланцюги; 7 — ферменти; 8
— амінокисло-ти;
9—
транспортна РНК (тРНК)
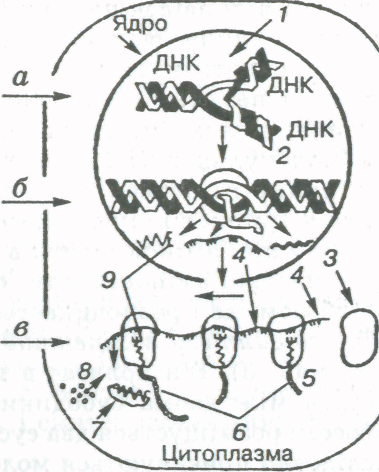 Інформація
про
структуру конкретного білка (наприклад,
якогось ферменту)закодована в молекулі
ДНК, яка міститься в ядрі. Ця інформація
може бути переписана на молекулу іРНК
лише в
Інформація
про
структуру конкретного білка (наприклад,
якогось ферменту)закодована в молекулі
ДНК, яка міститься в ядрі. Ця інформація
може бути переписана на молекулу іРНК
лише в
тому разі, коли подвійний ланцюг ДНК на певному відрізку роз'єднається і кожний із ланцюгів відійде один від одного. Цей процес здійснюється за допомогою специфічних ферментів, які розривають водневі зв'язки між азотистими основами окремих ланцюгів. Далі за участю ферменту РНК-полімерази вздовж одного із роз'єднаних ланцюгів ДНК розпочинається синтез молекули ІРНК. За принципом комплементарності послідовність нуклеотидів у ній повністю відповідатиме послідовності нуклеотидів в одному з ланцюгів молекули ДНК.
Як у друкарні з матриці можна видрукувати сотні тисяч примірників книг чи газет, так і з молекули ДНК можна отримати безліч точних копій у вигляді молекул іРНК. Отже, певна ділянка ДНК (ген) є матрицею для відповідної ІРНК.
Синтезовані молекули іРНК за участю інших ферментів відокремлюються від ланцюга ДНК і переходять із ядра в цитоплазму, де сполучаються з рибосомами ЕПС, а молекула ДНК відновлює свою структуру. Розміри молекули і РНК залежать від обсягу інформації про розміри молекули білка, закодованої в ній. Чим довша молекула іРНК, тим більша білкова молекула. Максимальна молекулярна маса і РНК може досягати 2 млн у. о.
II етап — активування амінокислот. Цей процес відбувається в цитоплазмі шляхом з'єднання різноманітних амінокислот, які утворюються під час розщеплення білків, із специфічними ферментами і молекулами АТФ. Активовані молекули амінокислот сполучаються з молекулами транспортної РНК (тРНК). Кожній з 20 відомих амінокислот відповідає певна тРНК. Усі вони мають невелику молекулярну масу, складаються з 70—80 нуклеотидів, які утворюють невеликі ланцюги у вигляді структур з кількома петлями. В петлях можуть виникати комплементарні зв'язки між нуклеотидами.
У молекулах тРНК є дві важливі ділянки: до однієї з них прикріплюється відповідна амінокислота, а інша містить триплет нуклеотидів (антикодон), який відповідає коду цієї амінокислоти в молекулі іРНК.
Активовані амінокислоти, сполучені з тРНК, надходять до рибосом, де і розпочинається Ш етап — процес безпосереднього синтезу поліпептидних ланцюгів — трансляція (див. мал. 5). Він полягає в тому, що молекула іРНК рухається між двома субодиницями рибосом (одночасно в рибосомі розміщується два сусідніх триплети іРНК) і до неї послідовно приєднуються молекули тРНК з активованими амінокислотами. При цьому рибосома за допомогою свого функціонального центру розпізнає антикодон тРНК і звільняє від неї амінокислоту, збираючи поліпептидний ланцюг.
Послідовність розміщення амінокислот визначається порядком чергування триплетів у молекулі іРНК. Опинившись поряд, амінокислоти утворюють пептидні зв'язки одна з одною, використовуючи енергію АТФ. У результаті з рибосоми сходить поліпептидний ланцюг (первинна структура білка), який містить багато амінокислотних залишків, розміщених у певній послідовності. Швидкість сполучення амінокислот між собою досягає 20—50 за секунду. Тому синтез поліпептидного ланцюга з 150 амінокислот відбувається за 3—5 с. Як правило, на іРНК під час біосинтезу знаходиться по кілька рибосом. У цьому разі в кожній окремій рибосомі, як на матриці, синтезуються однакові поліпептидні ланцюги (матричний синтез). IV етап — утворення вторинної і третинної структур білкової моле/сули. Цей етап відбувається в ЕПС шляхом скручування, згортання поліпептидного ланцюга. Утворення зв'язків визначається властивостями кожної окремої амінокислоти в ланцюзі.
Авторегуляція хімічної активності клітин. Незважаючи на безперервні процеси розщеплення і синтезу в клітині, надходження і виділення різних хімічних сполук, вміст білків, нуклеїнових кислот, вуглеводів та інших речовин у цитоплазмі підтримується відносно сталим (див. "Хімічна організація клітини"). Ця сталість складу зберігається лише в живих клітинах, а в разі відмирання їх вона порушується дуже швидко. Стійкість клітин (як і інших жи-50
систем) підтримується активно внаслідок перебігу
ладних процесів саморегуляції, або авторегуляції. Найтростіше авторегуляція здійснюється за принципом зв-ротного зв'язку. Він полягає у тому, що сама хімічна речовина (наприклад, білок), яка нагромаджується в клітині в результаті синтезу, гальмує свій подальший синтез, коли вміст її досягає певного рівня. Активність ферментів може регулюватися шляхом гальмування кінцевим продуктом. Наприклад, у клітинах амінокислота треонін внаслідок перебігу п'яти послідовних реакцій перетворюється на іншу амінокислоту — ізолейцин. Якщо в цитоплазмі нагромаджується достатня кількість Ізолейцину, то відбувається пригнічення активності ферменту, від якого залежить перша з п'яти реакцій, і синтез ізолейцину припиняється.
Другий регуляторний механізм хімічних процесів у клітині полягає у зміні активності ферментів внаслідок приєднання або відщеплення від їхніх молекул деяких сполук — сульфгідрильних груп (—SН), залишків фосфорної кислоти, аденозину.
Такі модифіковані ферменти стають активними або, навпаки, неактивними і відповідно "включаються" в хімічні процеси або "виключаються" з них. Ці хімічні модифікації здійснюються регуляторними ферментами, одні з яких активують, а інші інактивують молекули основних ферментів.
Ще один механізм регулювання хімічних процесів полягає в тому, що синтез певних ферментів у клітині розпочинається лише в тому разі, коли в неї потрапляють речовини, для розщеплення яких потрібні ці ферменти. Наприклад, зазвичай у клітинах дріжджів за участю багатьох ферментів відбувається розщеплення глюкози. Проте якщо в ці клітини потрапляє лактоза, то через деякий час у них синтезуються нові ферменти, які забезпечують можливість використання цього вуглеводу. Синтез ферментів зумовлений "включенням у роботу" певних генів, які раніше не функціонували. Отже, регуляція хімічних процесів пов'язана з пригніченням або стимулюванням певних генів.
Контрольні запитання і завдання
1. Дайте визначення поняття "обмін речовин" та схарактеризуйте його роль у житті організмів.
2. Що називають пластичним і енергетичним обміном?
3. Яка роль АТФ в обміні речовин?
4. Схарактеризуйте етапи енергетичного обміну.
5. Порівняйте процеси гліколізу і дихання за їх енергетичними показниками.
6. Які організми називають автотрофними, а які гетеротрофними І чому?
7. Дайте характеристику світлової І темнової стадій фотосинтезу.
8. Яке значення має фотосинтез у колообігу речовин?
9. Якими методами можна підвищити продуктивність рослин?
10. Дайте характеристику матричного синтезу в клітині.
11. Дайте визначення поняття "ген".
12. Що таке код ДНК, кодони та антикодони?
13. Назвіть основні етапи біосинтезу білка.
14. Яка роль нуклеїнових кислот (ДНК, іРНК, рРНК, тРНК) у біосинтезі білків?
16. Що таке транскрипція, трансляція?
16, Яка роль ферментів у процесах обміну речовин?
17. Які особливості будови І життєдіяльності вірусів?