

Энергетический обмен
Энергетический обмен в клетке подразделяют на три этапа. Первый этап – подготовительный. Во время него крупные пищевые полимерные молекулы распадаются на мелкие (мономеры). Главным поставщиком энергии для АТФ являются углеводы. Полисахариды (крахмал, гликоген) на подготовительном этапе расщепляются до глюкозы.
Процессы подготовительного этапа протекают в лизосомах, энергия при этом рассеивается в виде тепла, АТФ не образуется.
Второй этап – неполное бескислородное расщепление глюкозы – гликолиз – протекает в цитоплазме при помощи большого количества ферментов. В результате 1 молекула глюкозы расщепляется на 2 молекулы пировиноградной кислоты (ПВК), при этом выделяется энергия, достаточная, чтобы синтезировать 2 молекулы АТФ. Затем ПВК транспортируется в митохондрии, где протекает третий, заключительный этап энергетического обмена – клеточное дыхание.
Однако, при недостаточном количестве кислорода третий этап протекать не сможет. В таком случае в клетках животных и человека образовавшаяся ПВК превращается в молочную кислоту (молочнокислое брожение). А в клетках некоторых грибов, например, дрожжей, ПВК превращается в этиловый спирт и углекислый газ (спиртовой брожение).
Третий этап энергетического обмена – полное кислородное (аэробное) расщепление, или клеточное дыхание. На этом этапе ПВК, образовавшаяся при гликолизе, разрушается до углекислого газа и воды. Этот процесс протекает в митохондриях. В нем принимают участие многочисленные ферменты, молекулы-переносчики, вода, молекулярный кислород (О2). Основным условием нормального течения кислородного расщепления является целостность мембран митохондрий, в которые встроены молекулы цепи переноса электронов. На третьем этапе протекают такие сложные процессы, как окислительное декарбоксилирование, цикл трикарбоновых кислот (цикл Кребса), электрон-транспортная цепь. В результате образуется углекислый газ, вода, а энергии выделяется столько, что хватает на синтез 36 молекул АТФ (с 2 молекул ПВК)
Таким образом, суммарное уравнение энергетического обмена (подготовительный этап + гликолиз клеточное дыхание) выглядит так:
С6Н12О6 + 6 О2 +38АДФ + 38Н3РО4 38АТФ + 6СО2 + 44Н2О.
Матричные процессы.
Матричными называются такие процессы биосинтеза, в ходе которых на основе первичной структуры одного биополимера (матрицы) синтезируется первичная структура другого биополимера (копии); при этом структура матрицы однозначно определяет структуру копии.
В живой природе существует 3 фундаментальных матричных процесса: репликация ДНК, биосинтез РНК (транскрипция) и биосинтез белка (трансляция).
Общие черты матричных процессов:
идут согласно принципу комплементарности
требуют больших затрат энергии АТФ (или ГТФ)
все требуют участия ферментов и неферментных белковых факторов
включают три стадии: инициация, элонгация и терминация.
Биосинтез белка.
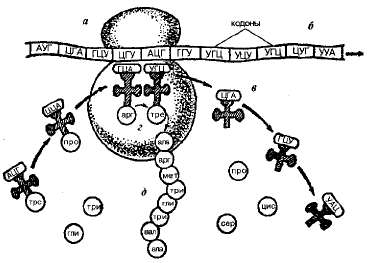
Ген - участок молекулы ДНК, содержащий информацию о первичной структуре одного белка. ДНК состоит из нуклеотидов (А,Г,Т,Ц), а белки построены из 20 разных аминокислот, соединенных в определенном порядке в последовательность из 300-500 мономеров. Зависимость между последовательностью нуклеотидов в молекуле ДНК и последовательностью аминокислот в молекуле белка называется генетическим кодом.
Главные черты генетического кода можно сформулировать следующим образом:
Кодом, определяющим включение аминокислоты в полипептидную цепь, служит триплет оснований в полинуклеотидной цепи ДНК. Код триплетен.
Код неперекрывающийся. Ни один нуклеотид одного триплета не является частью другого триплета (жил был дед)
Код универсален: одни и те же триплеты кодируют одни и те же аминокислоты у всех организмов на Земле (о единстве всего живого на планете).
Код однозначен (специфичен): один триплет шифрует только одну аминокислоту.
Код является вырожденным (избыточным): данная аминокислота может кодироваться более чем одним триплетом.
Некоторые последовательности являются знаками препинания (АТТ, АТЦ, АЦТ).
Транскрипция – матричный процесс, в ходе которого на матрице одной цепи ДНК (смысловой, или кодогенной) синтезируется копия – молекула РНК (по правилу комплементарности, только напротив аденина ДНК встает урацил РНК); осуществляется РНК-полимеразой, требует затрат энергии. Напротив А (ДНК) встает У (в РНК), напротив Т - А, напротив Г - Ц, напротив Ц - Г.
Образовавшаяся в результате транскрипции и-РНК покидает ядро через ядерные поры и выходит в цитоплазму, направляясь к шероховатой ЭПС.
Трансляция – такой матричный процесс, в ходе которого на матрице иРНК синтезируется копия – полипептид. Таким образом, информация о последовательности аминокислот в полипептиде содержится в последовательности нуклеотидов матричной РНК. Трансляция обеспечивается наличием белоксинтезирующего аппарата (рибосомы, тРНК, иРНК), для этого необходима энергия АТФ.
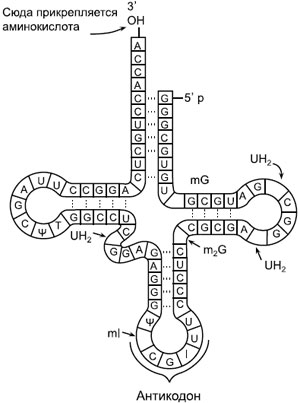
Рибосомы рассеяны в цитоплазме в неактивном виде (субъединицы). При объединении большой и малой субъединиц рибосома приходит в рабочее состояние (образуется 3 активных центра). В функциональном центре рибосомы могут одновременно находиться 6 нуклеотидов иРНК (2 триплета, 2 кодона), соответствующие двум антикодонам молекул тРНК. тРНК обеспечивают связывание и перенос молекул аминокислот в рибосому. Каждой аминокислоте соответствует одна или несколько тРНК. Информация о том, какую именно аминокислоту переносит тРНК, можно прочесть в антикодоне, комплементарном кодону иРНК. Аминокислота присоединяется к акцепторному концу молекулы - «ножке клевера» - при помощи особого фермента. Инициация цепи всегда начинается с аминокислоты метионина, соседний кодон может быть любым. Метионин присоединяется к соседней аминокислоте пептидной связью, тРНК отваливается, освобождая место, и рибосома делает «шажок» на 1 триплет. На свободное место приходит следующая тРНК с новой аминокислотой. Следующий этап – элонгация – удлинение цепи. |
|
Чем дальше рибосома продвинулась по и-РНК, тем больший кусок молекулы белка собран.
|
|
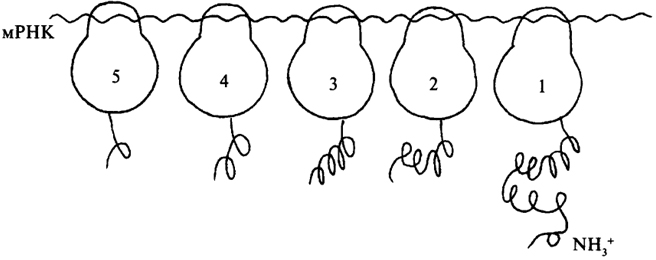 Несколько рибосом
могут прикрепиться к общей молекуле
и-РНК, образуя структуру, называемую
полисомой.
Преимущество такого комплекса в том,
что при этом на одной молекуле иРНК
становится возможным одновременный
синтез нескольких полипептидных
цепей. Все рибосомы полисомы синтезирует
один и тот же белок. Сборка полипептидной
цепи осуществляется с большой скоростью:
белок из 300 аминокислот синтезируется
Несколько рибосом
могут прикрепиться к общей молекуле
и-РНК, образуя структуру, называемую
полисомой.
Преимущество такого комплекса в том,
что при этом на одной молекуле иРНК
становится возможным одновременный
синтез нескольких полипептидных
цепей. Все рибосомы полисомы синтезирует
один и тот же белок. Сборка полипептидной
цепи осуществляется с большой скоростью:
белок из 300 аминокислот синтезируетсяза минуту.

Когда синтез окончен, рибосома сходит с цепи и-РНК. Дальше она может остаться в цитоплазме или присоединиться к любой другой молекуле и-РНК, т.к. рибосома способна синтезировать любой белок, а какой, это написано в и-РНК, к которой она присоединится. Белок поступает в ЭПс и транспортируется в заданном направлении.
В клетках содержится множество генов. Но в каждый момент жизни клетки в ней работают только те гены, в которых есть необходимость, т.е. в разных клетках в разное время транскрибируются разные участки ДНК и образуются разные иРНК, по которым синтезируются разные белки. Имеется сложный механизм, регулирующий «включение» и «выключение» генов на разных этапах жизни клетки.
ДОПОЛНИТЕЛЬНЫЙ МАТЕРИАЛ
Если для примера взять относительно просто устроенную бактериальную клетку, получится следующая картинка. Известно, что, пока в питательную среду, в которой живет бактерия, не добавлен сахар, в клетке нет ферментов, необходимых для его расщепления. Бактерия не тратит драгоценную АТФ на синтез белков, ненужных ей в данный момент, все равно они быстро денатурируют. Однако через несколько секунд после добавления сахара в клетке синтезируются все ферменты, последовательно превращающие его в продукт, необходимый для жизнедеятельности бактерий. Вместо сахара может быть другое соединение (субстрат), появление которого в клетке «включает» синтез ферментов, расщепляющих его до конечного продукта.
Единица транскрипции прокариот называется оперон. Концепция оперона разработана Ф.Жакобом и Ж.Моно в 1961. Оперон состоит из нескольких структурных генов, расположенных последовательно друг за другом, промотора и терминатора. Между структурными генами и промотором имеется особый участок ДНК – оператор, с которым взаимодействует специальный белок – репрессор. Пока репрессор сидит на операторе, РНК-полимераза не может сдвинуться с места.
Когда в клетку попадает субстрат А, для расщепления которого нужны ферменты Ф1, Ф2, Ф3, закодированные в структурных генах 1,2,3 оперона А, одна из молекул субстрата связывается с репрессором, в результате чего последний теряет связь с матрицей. Путь для РНК-полимеразы освобождается, и она синтезирует иРНК, содержащую информацию о трех соответствующих белках. Такие иРНК называются полицистронными, и характерны для прокариот. Белки, кодируемые полицистронными иРНК, обычно функционально связаны друг с другом и обеспечивают протекание какого-либо метаболического процесса. Как только последняя молекула субстрата А будет переработана ферментами, репрессор освободится и снова свяжется с оператором, и транскрипция прекратится.
Организация генов в виде оперонов облегчает координированную регуляцию их экспрессии на уровне транскрипции.
У эукариот в состав транскриптона, как правило, входит только один ген. Соседние транскриптоны могут быть отделены друг от друга нетранскрибируемыми участками ДНК, а могут и перекрываться, в частности так, что в пределах участка перекрывания матричными оказываются обе нити. Разбиение ДНК на множество транскриптонов обеспечивает возможность независимого считывания разных генов, их индивидуального включения и выключения.
Согласованная регуляция транскрипции (и других этапов экспрессии) многих генов, не образующих одного оперона, чаще всего осуществляется специфическими белками-регуляторами, которые взаимодействуют с гомологичными регуляторными нуклеотидными последовательностями, маркирующими гены данной группы.