


2. РЕПЛИКАЦИЯ, СОХРАНЕНИЕ И МОДИФИКАЦИЯ ГЕНОМА |
103 |
при очень слабом воздействии солнечного света. Клетки таких людей несут мутацию, сходную с RAD-мутацией дрожжей и проявляющуюся в том, что у них нарушена способность к выщеплению пиримидиновых димеров из УФ-облученной ДНК. Заболевание может быть обусловлено мутацией в одном из по крайней мере девяти генов, что говорит о достаточно сложном механизме репарации ДНК, содержащей тиминовые димеры, у человека. Как правило, заболевание бывает связано с неспособностью к выщеплению тиминовых димеров. Если к облученным клеткам в культуре добавить фермент, обладающий тиминдимергликозилазной и АР-эндонуклеазной активностями, то УФ-повреж- дения могут быть устранены.
2.4. РЕКОМБИНАЦИЯ ДНК
Генетическая рекомбинация включает несколько связанных между собой процессов, в результате которых в клетках или организмах, где они происходят, создаются новые комбинации элементов-
носителей генетической информации. Рекомбинация между близко расположенными гомологичными хромосомами приводит к интенсивной перетасовке отцовских и материнских генов в ходе мейоза и тем самым создает предпосылки для эволюционной проверки новых комбинаций этих генов в потомстве. Как правило, рекомбинационные события, происходящие в соматических клетках либо во время репликации ДНК, либо после нее и проявляющиеся в виде обмена сестринских хроматид (рис. 2.51), не приводят к изменению генотипа или фенотипа клетки. Однако нередко они порождают различные геномные перестройки. Это, например, утрата, приобретение или амплификация генетических элементов и установление новых взаимосвязей между уже имеющимися, но по-новому расположенными элементами.
Если использовать молекулярные термины, то можно сказать, что генетическая рекомбинация состоит в образовании ковалентных связей между нуклеотидными последовательностями из разных областей одной и той же или разных молекул ДНК.
А Б
РИС. 2.51.
Обмен сестринских хроматид. Если использовать спе- |
сестринские хроматиды, образовавшиеся во втором |
||
циальную методику, то можно увидеть, как в метафаз- |
клеточном цикле, имеют более интенсивную окраску. |
||
ных хромосомах происходит обмен между двумя про- |
Обмен сестринских хроматид выявляется по неравно- |
||
дуктами дупликации хромосом (сестринскими хрома- |
мерности интенсивности флуоресценции. А. Хромосо- |
||
тидами). Методика вкратце состоит в следующем: клет- |
мы из нормальных периферических лимфоцитов че- |
||
ки проходят одно деление в присутствии бромдезокси- |
ловека. Б. Хромосомы из периферических лимфоцитов |
||
уридина, а второе - в его отсутствие. Краситель Hoechst |
больных |
с синдромом Блума. |
[S.A. Latt, R. Schreck, |
33258 в комплексе с ДНК, содержащей бромдезокси- |
Amer. J. Hum. Genet., 32 (1980), p. 294.] Микрофо- |
||
уридин, флуоресцирует значительно слабее, чем в |
тографии |
любезно предоставлены |
S.A. Latt. |
комплексе с обычной тиминсодержащей ДНК. Поэтому |
|
|
|

104 |
ЧАСТЬ I. МОЛЕКУЛЯРНЫЕ ОСНОВЫ НАСЛЕДСТВЕННОСТИ: ОБЗОР |
Все клетки и многие вирусы содержат информацию о синтезе ферментов, предназначенных не только для репарации повреждений в собственной ДНК, но и ферментов, осуществляющих рекомбинацию. На самом деле некоторые ферменты, участвующие в репликации и репарации ДНК, играют ключевую роль и при рекомбинации. В этом разделе мы рассмотрим механизмы некоторых рекомбинационных процессов и ферменты, которые их катализируют. Особое внимание будет обращено на рекомбинацию у бактерий и фагов, поскольку у них эти процессы довольно хорошо изучены. Несмотря на то что генетические и морфологические аспекты рекомбинации в эукариотических клетках известны, на молекулярном уровне здесь многое остается неясным.
а. Типы рекомбинации
Существуют три типа рекомбинации: общая, или
гомологичная, сайт-специфическая и случайная, или негомологичная.
Общая рекомбинация. Общая рекомбинация происходит, как правило, между протяженными участками идентичных или гомологичных нуклеотидных последовательностей. Ее часто называют гомологичной рекомбинацией или кроссинговером. При общей рекомбинации происходит разрыв двух гомологичных участков ДНК, и каждый из концов одного сегмента соединяется с соответствующими концами другого таким образом, что обе образующиеся молекулы содержат разные фрагменты обеих участвующих в рекомбинации ДНК (рис. 2.52). На самом деле сайты, по которым происходят разрыв и воссоединение каждой из двух цепей, очень часто не совпадают.
Обычно общая рекомбинация происходит между гомологичными и аллельными участками разных молекул ДНК, но она может произойти и между гомологичными, но неаллельными областями рекомбинирующих молекул. В этом случае один из продуктов рекомбинации утрачивает часть ДНК,
РИС. 2.52.
Гомологичная рекомбинация, или кроссинговер.
РИС. 2.53.
Неравный кроссинговер.
а другой приобретает «лишний» сегмент. Такой процесс получил название неравного кроссинговера (рис. 2.53). Иногда рекомбинация происходит между неаллельными участками одной и той же хромосомы (например, между повторяющимися последовательностями ДНК) с соответствующей потерей области, лежащей между сайтами рекомбинации (рис. 2.54). В отличие от уже рассмотренных случаев некоторые рекомбинационные события нереципрокны; как следствие, один из образовавшихся продуктов идентичен одной из исходных молекул, а другой отличается от обоих партнеров (рис. 2.55). Такой процесс часто называется генной конверсией.
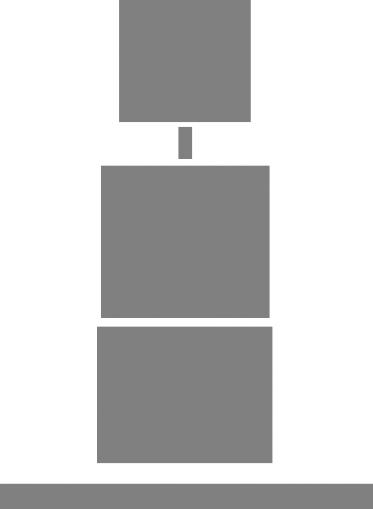
Сайт-специфическая рекомбинация. Рекомбинация называется сайт-специфической, если сайты разрыва и воссоединения в двух рекомбинирующих молекулах или двух фрагментах одной и той же молекулы ДНК находятся в пределах довольно коротких специфических гомологичных нуклеотидных последовательностей–как правило, не более 25 нуклеотидов. Такие короткие последовательности может иметь только один из партнеров (рис. 2.56) или оба (рис. 2.57). В качестве примера первого варианта можно привести транспозиции некоторых мобильных элементов у эу- и прокариот (разд. 10.2 и 10.3), а второго–процесс интеграции-выщепления ДНК фага λ из хромосомы Е. coli (разд. 2.4.г). С помощью сайт-специфической рекомбинации происходят запрограммированные перестройки хромосомной ДНК при смене типов спаривания у дрожжей; она ответственна также за разнообразие антител (разд. 10.6). По-видимому, общая рекомбинация между любыми парами гомологичных последовательностей осуществляется с помощью одного и того же комплекса ферментов; с другой стороны, для каждого случая сайт-специфической рекомбинации необходим свой набор ферментов.
2. РЕПЛИКАЦИЯ, СОХРАНЕНИЕ И МОДИФИКАЦИЯ ГЕНОМА |
105 |
Рекомбинация |
+ |
+
РИС. 2.54.
Внутримолекулярная рекомбинация.
Негомологичная рекомбинация. Рекомбинация между негомологичными нуклеотидными последовательностями происходит в клетках прокариот и дрожжей достаточно редко, а в клетках млекопи- тающих–весьма часто. К негомологичной рекомбинации можно отнести процесс случайного встраивания вирусной или плазмидной ДНК в ДНК клеток животных, в результате чего в реплицирующихся геномах паповавирусов появляется множество делеций и дупликаций. Концы разорванной ДНК могут соединиться, даже если они негомологичны. В некоторых случаях рекомбинация происходит между последовательностями, содержащими несколько гомологичных пар оснований, или между короткими
Генная конверсия
РИС. 2.55.
Нереципрокная рекомбинация, или генная конверсия.
Рис. 2.56.
Сайт-специфическая рекомбинация, при которой специфический сайт имеется только в одном участвующем в рекомбинации фрагменте ДНК. Специфическими сайтами рекомбинации являются концы участка Т, а сам участок может рекомбинировать со случайными последовательностями ДНК, расположенными, например, между участками В и С (вверху) или в пределах участка D (внизу).
частично гомологичными участками. Но, как правило, рекомбинирующие сегменты не имеют гомологичных последовательностей.
б. Общая рекомбинация между гомологичными молекулами ДНК
Общая рекомбинация при согласованном внесении разрывов и воссоединении цепей двух спиралей ДНК с образованием протяженных гетеродуплексных областей. Чтобы могла произойти рекомбинация между двойными спиралями, представленная на рис. 2.52, каждая из четырех цепей должна быть разорвана и затем соединена с новым партнером. Этапы этого процесса показаны на рис. 2.58. Соответствующие цепи обоих линейных гомологичных дуплексов ДНК надрезаются и свободные концы одной спирали спариваются с комплементарными участками другой (а-г). Перекрест стабилизируется сшиванием концов донорных цепей со свободными концами реципиентных спиралей (д). Точка перекреста обменивающихся цепей перемещается вдоль
106 |
ЧАСТЬ I. МОЛЕКУЛЯРНЫЕ ОСНОВЫ НАСЛЕДСТВЕННОСТИ: ОБЗОР |
РИС.2.57.
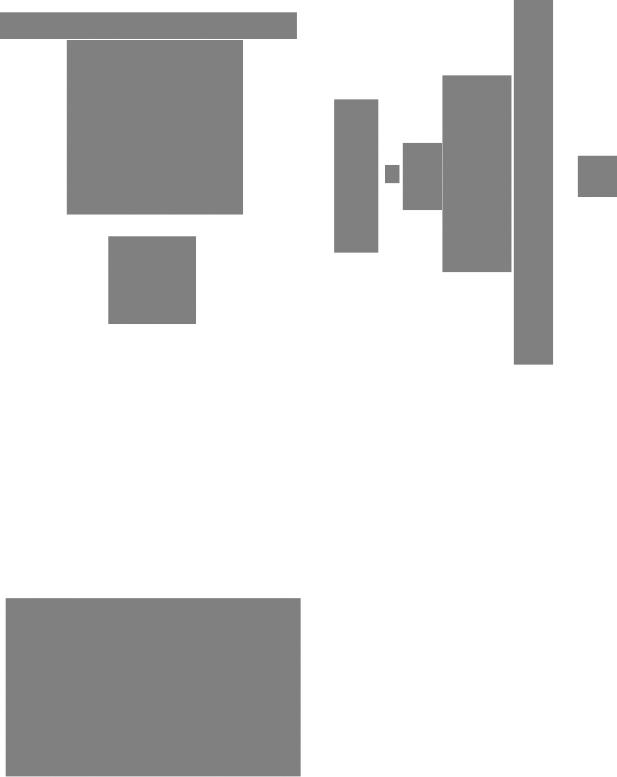
Сайт-специфическая рекомбинация, при которой специфические последовательности имеются в обеих участвующих в рекомбинации ДНК. На рисунке представлена интеграция ДНК фага λ с хромосомной ДНК Е. соli. [В. Alberts et al.. Molecular Biology of the Cell (New York: Garland, 1983), p. 248.]
Геном бактериофага λ (кольцевая
двухцепочечная ДНК)
Специфические
последовательности, по которым
происходит встраивание Хромосомная ДНК
Белковый комплекс катализируетсогласованное
образование двухцепочечного разрыва и
воссоединение дуплексов
Белок
ДНК бактериофага встроилась в хромосомную ДНК
спиралей – процесс, называемый миграцией ветви (е). При этом происходит одновременное расхождение цепей исходных спиралей и их реассоциация с новыми партнерами с образованием дочерних дуплексов. Структуры д и е, а также ж называются структурами Холлидея по имени исследователя, впервые их предложившего.
Структуры Холлидея могут переходить в рекомбинантные двойные спирали путем внесения разрыва и воссоединения цепей двумя альтернативными способами. Один способ состоит в разрезании и воссоединении перекрещивающихся цепей. Два реципрокных продукта л и м могут образоваться, если разрыв и последующее воссоединение цепей произойдут в точке перекреста в структурах е и д или по линии пересечения четырех цепей в изомерной структуре Холлидея и. Размер обменивающихся фрагментов зависит от расстояния, на которое про-
изошла миграция ветви до акта рекомбинации. Альтернативные продукты н и о образуются в том случае, если структура Холлидея з переходит в результате разрыва в к.
В основе рекомбинации данного типа лежит гомологичное спаривание цепей, принадлежащих двум разным спиралям ДНК, поэтому скорее всего она произойдет в том месте, где такое спаривание возможно a priori и где гомологичность последовательностей достаточно велика, чтобы могла произойти миграция ветви в рамках структуры со скрестившимися цепями. Отсюда можно понять, почему общая, или гомологичная, рекомбинация происходит также между двумя повторами в пределах одной молекулы ДНК или между аллельными и неаллельными элементами одной и той же последовательности в двух разных хромосомах (рис. 2.53 и 2.54).
В ходе миграции ветви при спаривании цепей,

2. РЕПЛИКАЦИЯ, СОХРАНЕНИЕ И МОДИФИКАЦИЯ ГЕНОМА |
107 |
РИС. 2.58.
Этапы процесса рекомбинации между гомологичными дуплексными ДНК (объяснения см. в тексте).
принадлежащих разным спиралям, образуются гете- |
ваний, т.е. рекомбинация для этого сайта окажется |
родуплексы. В таких гетеродуплексах в пределах |
нереципрокной (рис. 2.59). Таким образом, каждая |
сегмента между сайтом начала образования струк- |
из рекомбинантных спиралей может быть похожа на |
туры Холлидея и сайтом кроссинговера может со- |
любой из начальных дуплексов в тех позициях, где |
держаться по одному или более ошибочно спарен- |
исходно они различались. |
ных оснований (рис. 2.58). Они удаляются так же, |
Общая рекомбинация с образованием двухцепочеч- |
как любые модифицированные основания при репа- |
ного разрыва. Альтернативный механизм общей ре- |
рации ДНК (разд. 2.3.б). Однако, поскольку удале- |
комбинации включает образование двухцепочечно- |
но может быть любое из ошибочно спаренных |
го разрыва в одном из дуплексов-партнеров (рис. |
оснований, в обеих рекомбинантных спиралях в дан- |
2.60). Далее с помощью экзонуклеаз в месте разрыва |
ном сайте могут оказаться одинаковые пары осно- |
образуется брешь. При спаривании 3'-одноцепочеч- |

108 |
ЧАСТЬ I. МОЛЕКУЛЯРНЫЕ ОСНОВЫ НАСЛЕДСТВЕННОСТИ: ОБЗОР |
РИС. 2.59.
Продукты, образующиеся при неправильном спаривании в процессе гомологичной рекомбинации при репарации гетеродуплексов.
+
Гетеродуплексы, образующиеся из структур Холлидея
Исправление ошибок, происходящих при спаривании
или |
или |
ного конца бреши с комплементарной цепью ин- |
вательность одного дуплекса замещается соответст- |
||
тактной спирали в последней образуется петля. |
вующей, |
но отличающейся |
последовательностью |
Размер этой петли увеличивается по мере того, как |
другого |
дуплекса. |
|
ДНК-полимераза наращивает 3'-конец «вклинив- |
Нереципрокная общая рекомбинация использу- |
||
шейся» цепи. В итоге другой одноцепочечный конец |
ется и при репарации некоторых повреждений ДНК. |
||
бреши спаривается с комплементарной последова- |
Например, если тиминовые димеры не были удале- |
||
тельностью в перемещающейся петле. В результате |
ны из УФ-облученной ДНК до того, как к ним |
||
такого спаривания образуется система «праймер- |
подошла репликативная вилка, то синтез компле- |
||
матрица», и ДНК-полимераза синтезирует недоста- |
ментарной цепи в этом участке не может быть |
||
ющую цепь, заполняя брешь. Лигирование двух |
завершен (рис. 2.61). Поскольку тиминовые димеры, |
||
растущих концов с исходными цепями приводит |
находящиеся напротив бреши, не могут быть вы- |
||
к образованию двойной структуры Холлидея (т.е. |
щеплены, остается один путь для спасения хрома- |
||
структуры, в которой две спирали объединены дву- |
тиды–использовать генетическую информацию го- |
||
мя перекрестами, по одному на каждом конце бре- |
мологичной сестринской хроматиды и заполнить |
||
ши). Миграция ветви в одном или обоих перекрес- |
брешь. Для этого применяется такой же механизм, |
||
тах передвигает оба места сцепления в любом на- |
как для репарации брешей (рис. 2.60). |
||
правлении, при этом в участках, фланкирующих |
|
|
|
брешь, могут возникать ошибки. Разделение таких |
|
в. Ферменты, |
|
структур может идти двумя способами–с перекрес- |
участвующие в общей рекомбинации |
||
том и без него (рис. 2.58), с образованием четырех |
|
|
|
дуплексов (рис. 2.60). |
В общей рекомбинации участвуют два специфи- |
||
Необходимо отметить некоторые особенности |
ческих фермента и еще несколько ферментов, ката- |
||
этого механизма. Образование ошибочных пар (ге- |
лизирующих также процессы репликации и репара- |
||
теродуплексов) в районах, фланкирующих брешь, |
ции ДНК. Энзимология общей рекомбинации изуче- |
||
обусловливает получение как реципрокных, так и не- |
на только для некоторых прокариотических орга- |
||
реципрокных рекомбинаций между генетическими |
низмов, |
в частности E. coli |
и ее фагов. Один из |
маркерами. Если двухцепочечный разрыв происхо- |
специфических ферментов, необходимых для успеш- |
||
дит вблизи (или в пределах) участка, где между |
ной гомологичной рекомбинации, называется recА- |
||
спиралями имеются различия (замены оснований, |
белком. Он катализирует обмен одиночными цепя- |
||
делеции, вставки, инверсии и т.п.), то рекомбинан- |
ми, используя энергию гидролиза АТР до ADP |
||
ты унаследуют нуклеотидную последовательность |
и неорганического фосфата (рис. 2.62). RecA-зави- |
||
партнера, у которого разрыва не происходило. Этот |
симое внедрение одноцепочечных ДНК в дуплекс – |
||
механизм объясняет многие случаи генной конвер- |
первый этап рекомбинационного процесса в рамках |
||
сии, особенно те, в которых протяженная последо- |
обеих схем Холлидея (рис. 2.58) и механизма с обра- |
||

Без перекреста
С перекрестом
2. РЕПЛИКАЦИЯ, СОХРАНЕНИЕ И МОДИФИКАЦИЯ ГЕНОМА |
109 |
Двухцепочечный разрыв
 Экзонуклеазное расщепление
Экзонуклеазное расщепление
Образование петли
Увеличение размера петли с помощью Pol I
Репарация брешей и миграция ветви
Образованиеконечных продуктов
+
 или
или
+
РИС. 2.60.
Общая рекомбинация с образованием двухцепочечного разрыва.
зованием двухцепочечных разрывов (рис. 2.60). Второй фермент, состоящий из трех отдельных субъединиц (В, С и D) и поэтому называемый recBCD-нук- леазой, обладает эндо- и экзонуклеазной, а также геликазной активностями. Механизм его действия до конца не установлен, однако известно, что recBCD-нуклеаза индуцирует разрывы в дуплексной ДНК и благодаря присущей ей геликазной активности вместе с recА инициирует рекомбинационный
процесс. Идентифицирован также фермент, разрезающий узлы в структурах Холлидея; при его участии образуются липкие концы, соединяемые лигазой.
В общей рекомбинации участвуют также геликазы и белки, связывающиеся с одноцепочечной ДНК (SSB; от англ. single strand binding); оба они необходимы для обеспечения процесса миграции ветви. Как известно, перемещению цепей во время миграции ветви способствует Pol I, а в воссоединении
110 |
ЧАСТЬ I. МОЛЕКУЛЯРНЫЕ ОСНОВЫ НАСЛЕДСТВЕННОСТИ: ОБЗОР |
|
|
Пиримидиновый димер |
Репарированный сегмент |
|
|
Обычная |
|
|
репарация |
|
Репарация бреши |
пиримидинового |
|
|
димера |
Донорный сегмент, использующийся для репарации бреши
РИС.2.61.
Нереципрокная гомологичная рекомбинация как один из этапов репарации ДНК в месте образования пиримидиновых димеров.
разорванных цепей участвует ДНК-лигаза. Для снятия топологических ограничений при раскручивании спирали и для распутывания перекрученных структур, по-видимому, нужны топоизомераза типа I и, возможно, гираза.
|
|
г. Сайт-специфическая |
|
|
|
рекомбинация |
|
|
Сайт-специфическая рекомбинация происходит |
||
|
между специфическими сегментами дуплексов ДНК, |
||
|
не имеющими протяженных гомологичных участ- |
||
|
ков. Характерным примером такой рекомбинации |
||
|
служит интеграция кольцевой ДНК фага λ с хромо- |
||
|
сомой Е. coli и ее обратное выщепление. Несмотря |
||
|
на то что эти рекомбинационные события также |
||
RecA-белок, АТР |
включают разрыв и воссоединение двух спиральных |
||
SSB |
сегментов ДНК, их механизм абсолютно отличен от |
||
|
механизма общей рекомбинации. В этом случае |
||
|
рекомбинация происходит в пределах специфиче- |
||
|
ской нуклеотидной последовательности ДНК фага |
||
|
λ (attP-сайт) и уникальной последовательности |
||
|
ДНК |
Е. coli (аttВ-сайт) (рис. 2.63). Нуклеотидные |
|
SSB |
последовательности attP- и attВ-сайтов (рис. 2.64) |
||
|
совершенно различны, хотя имеют общее ядро (О) |
||
|
протяженностью в 15 нуклеотидных пар. AttP |
||
|
(POP') |
простирается на 150 нуклеотидов влево (Р) |
|
АТР |
и на |
75 нуклеотидов вправо (Р') от общего ядра, |
|
a attB (BOB')–это сегмент длиной всего около 25 |
|||
|
|||
RесА-белок, |
нуклеотидов, включая и ядро. Рекомбинационные |
||
SSB |
события, происходящие как при интеграции, так |
||
ADP + Pi |
и при исключении ДНК фага λ из хромосомы Е. coli, |
||
|
|||
|
изображены на рис. 2.63 (фрагменты ДНК представ- |
||
|
лены в виде линейных структур). |
||
|
Поскольку нуклеотидные последовательности, |
||
|
РИС. 2.62. |
||
|
RecA-зависимая общая рекомбинация ДНК E. coli. Про- |
||
|
цесс ускоряется присоединением белка, связывающего- |
||
Одноцепочечный |
ся с одноцепочечной ДНК (SSB). [M. Cox, I.R. Lehman, |
||
разрыв |
Proc. Natl. Acad. Sci. USA, 78 (1981), p. 3433.] |
||