

68) Методы изучения днк. Секвенирование генома. Современная геномика.
69)Онтогенез. Регуляция синтеза белка в клетке прокариотов по Жакобу и Моно
Онтогене́з (от греч. οντογένεση: ον — существо и γένεση — происхождение, рождение) — индивидуальное развитие организма отоплодотворения (при половом размножении) или от момента отделения от материнской особи (при бесполом размножении) до смерти.
Онтогенез – это индивидуальное развитие организма от момента его зарождения до смерти. Онтогенез начинается с оплодотворения (слияния сперматозоида и яйцеклетки). При этом образуется зигота, в которой объединяется наследственный материал отца и матери. Онтогенез состоит из 2 периодов:
1. эмбриональный - это период с момента оплодотворения до рождения. Рождение- это выход зародыша из утробы матери или яйцевых оболочек.
2. Постэмбриональный период- с момента рождения до момента смерти (прекращение всех жизненных процессов).

1. Зародышевое (эмбриональное) развитие
Включает в себя стадии дробления, гаструляции, органогенеза.
Дробление – это серия делений зиготы путем митоза на 2,4, 8, и т.д. клеток. Промежуток между делениями очень короткий, в нем происходит только удвоение ДНК, а рост клеток не происходит (яйцеклетка и так была очень большая). В процессе дробления клетки постепенно уменьшаются, пока не достигают нормальных размеров. После дробления образуется бластула – многоклеточный полый шарик из одного слоя мелких клеток - бластомеров.Внутри бластулы есть первичная полость- бластоцель.
Затем одна стенка бластулы начинает активно делиться и стенка впячивается во внутрь бластулы. Так бластула превращается вгаструлу – двухслойный мешок. Наружный слой клеток бластулы называется эктодерма, внутренний – энтодерма, отверстие в гаструле называется первичный рот, он ведет в кишечную полость. (Такое же, как у бластулы, строение имеют кишечнополостные, например, гидра.Это считается доказательством эволюции.). В конце гаструляции образуется третий зародышевый листок- мезодерма и прорывается вторичный рот на конце, противоположном первичному рту.
Органогенез (образование органов) начинается с формирования нервной пластинки в эктодерме на спинной стороне зародыша. нейрула- это стадия, на которой начинается образоваться нервная пластинка. В дальнейшем из трех зародышевых листков формируются:
из эктодермы образуется кожа и нервная система(головной и спинной мозг), органы чувств.
из энтодермы – пищеварительная (печень, поджелудочная железа и др.) и дыхательная система (трахеи, бронхи, легкие);
из мезодермы – всё остальное (скелет, мышцы, кровеносная, выделительная, половая системы).

2. Постэмбриональное развитие
Бывает двух видов:
Прямое – когда личинка похож на родителя, только меньше по размерам и у него недоразвиты некоторые органы (млекопитающие, птицы, рептили, из насекомых: тараканы, прямокрылые).
Непрямое (с превращением, с метаморфозом) – когда личинка сильно отличается от родителя по внешнему строению, питанию и образу жизни.например, головастик не похож на лягушку, а гусеница на бабочку. Преимущество непрямого развития состоит в том, что родители и дети не конкурируют друг с другом за пищу и территорию. Так развиваются: земноводные, чешуекрылые, жесткокрылые, блохи, двукрылые, перепончатокрылые). Стадии непрямого развития насекомых: яйцо, личинка, куколка, взрослая особь. Особенности жизни животных на стадии яйца и куколки — они неподвижны. Значение непрямого развития — ослабление конкуренции между родителями и потомством, так как они поедают разную пищу, у них разные места обитания. Непрямое развитие — важное приспособление, возникшее в процессе эволюции. Оно способствует ослаблению борьбы за существование между родителями и потомством, выживанию животных на ранних стадиях послезародышевого развития.
РАЗВИТИЕ ГОЛОВАСТИКА- ЭТО НЕПРЯМОЕ РАЗВИТИЕ С ПРЕВРАЩЕНИЕМ РАЗВИТИЕ ПРЯМОКРЫЛЫХ- ЭТО ПРИМЕР ПРЯМОГО РАЗВИТИЯ


Регуляция у прокариот
Регуляция биосинтеза белка упрокариот осуществляется на уровне изменения скорости синтеза мРНК. В настоящее время принята теория оперона, сформулированная Франсуа Жакобом и Жаком Моно. В основе теории лежат следующие понятия:
оперон – группа тесно связанных между собой генов, которые программируют образование структурных белков и ферментов в клетке,
конституитивные ферменты – те, которые присутствуют в клетках всегда, независимо от ее активности и условий,
индуцибельные ферменты – те, которые программируются опероном и синтезируются при необходимости,
ген-регулятор – ген, регулирующий работу оперона, но не входящий в его состав. Он синтезирует белок-регулятор (чаще называемый белок-репрессор), который может быть в активной или неактивной форме,
ген-оператор – участок ДНК, способный связываться с белком-регулятором, и "решающий" нужно работать РНК-полимеразе или нет.

Схема строения и работы оперона
Предложены две схемы регуляции скорости транскрипции: по механизму индукции(лактозный оперон) и по механизму репресии (триптофановый оперон).
Лактозный оперон
Лактозный оперон в целом отвечает за катаболизм лактозы.
При изучении E.coli было замечено, что в клетке может быть две взаимоисключающие ситуации:
активность одного из ферментов катаболизма лактозы низка, если в среде имеетсяглюкоза.
активность этого фермента резко повышается в обратной ситуации, т.е. при отсутствии глюкозы и при наличии лактозы.
На основании наблюдений была предложена схема регуляции оперона по механизму индукции:
1. В отсутствие лактозы активный белок-репрессор связывается с оператором и блокирует синтез мРНК, кодирующей ферменты катаболизма лактозы. В результате эти ферменты не образуются.
2. Если глюкозы нет, а лактоза есть, то последняя связывается с белком-репрессором и ингибирует его, не давая связаться с геном-оператором. Это позволяет РНК-полимеразе считывать информацию, отвечающую за синтез ферментов катаболизма лактозы, и синтезировать мРНК.
Таким образом, лактоза является индуктором транскрипции.
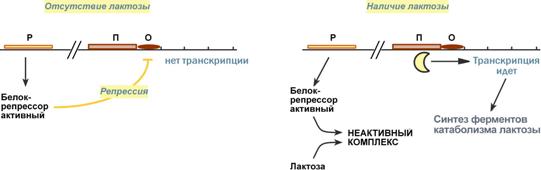
Схема работы лактозного оперона при наличии и отсутствии лактозы
Триптофановый оперон
Триптофановый оперон в целом отвечает за синтез триптофана.
Функционирование триптофанового оперона в некотором смысле противоположно лактозному. Регуляция осуществляется по механизму репрессии.
1. В отличие от лактозного оперона, белок-репрессор синтезируется в неактивномсостоянии и не может заблокировать транскрипцию генов, кодирующих ферменты синтеза триптофана. Синтез этой аминокислоты будет в клетке продолжаться до тех пор, пока в питательной среде не появится триптофан.
2. Триптофан соединяется с белком-репрессором и активирует его. Далее такой активный комплекс присоединяется к гену-оператору и блокирует транскрипцию. Таким образом, при наличии триптофана в среде прекращается его внутриклеточный синтез, экономятся ресурсы и энергия бактериальной клетки.
В этом случае триптофан является репрессором транскрипции.

Схема работы триптофанового оперона при наличии и отсутствии триптофана