

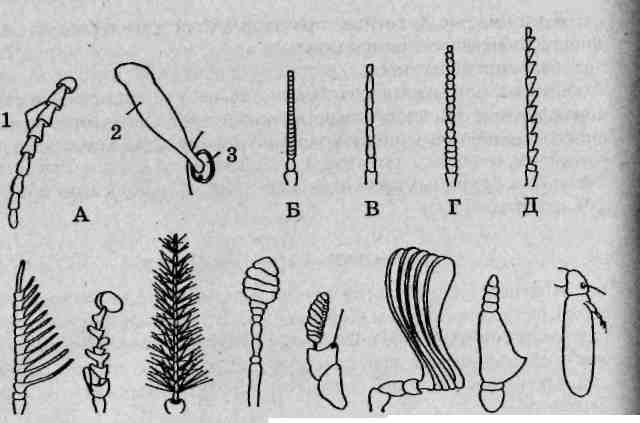
Особенности строения антенн
Антенны (усики) считаются придатками акрона (головной лопасти) и не гомологичны конечностям. Отсутствуют они только у бес-сяжковых (отряд Proturd). Основная функция антенн — чувствительная — обеспечивается наличием огромного числа рецепторов (химических, тактильных и др.). В составе многочлениковых антенн принято выделять:
основной членик (рукоятку),
ножку (педицелл), обычно имеющую наибольшую длину среди всех члеников,
— жгутик (бич), объединяющий все остальные членики (рис. 112). Строение антенн у насекомых разных эколого-систематических
групп весьма разнообразно. Для ознакомления с выделяемыми типами и особенностями строения антенн используются постоянные препараты, которые лучше просматривать под бинокуляром.
Нитевидные антенны состоят из многочисленных линейно расположенных члеников и имеют одинаковую толщину по всей своей длине.
Щетинковидные (шиловидные) антенны состоят из многочисленных линейно расположенных члеников и постепенно утончаются к вершине.
Веретеновидные антенны имеют наибольшую ширину в средней части и постепенно сужаются как к вершине, так и к основанию.
Четковидные антенны состоят из округлых или продолговато-овальных члеников, имеющих наименьший диаметр в местах при-членения к соседним.
Пиловидные (пильчатые) усики состоят из члеников, имеющих боковые выступы наподобие зубцов пилы.
220 Тип Arthropoda
Класс Insecta
221
жзикл мн о
Рис. 112. Антенны насекомых: А — общий план строения: 1 — жгутик; 2 — ножка; 3 — основной членик; Б — щетинковидная; В — нитевидная; Г— четковидная; Д— щитовидная; Е— булавовидная; I Ж— гребневидная; 3— неправильная; И— перистая; К— головчатая жука-мертвоеда; Л— головчатая блохи; М— пластинчатая; Я— неправильная с расчлененным члеником; О —щетинконосная
Гребневидные (гребенчатые) усики состоят из члеников, несущих с внешней стороны длинные боковые пластинчатые отростки, которые образуют своеобразную гребенку.
Перистые антенны состоят из члеников, несущих как с внешней, так и с внутренней стороны гибкие щетинковидные отростки, образующие характерное оперение.
Булавовидные усики отличаются утолщающимися вершинными члениками, образующими округлую компактную булаву.
Пластинчато-булавовидные усики иногда рассматриваются j в качестве варианта булавовидных. Вершинные членики сильно уко-рочены и несут крупные пластинчатые боковые отростки. Внутрен- ] няя мускулатура позволяет такой булаве веерообразно расправляться j и складываться.
Головчатые усики также рассматриваются как вариант булаво- | видных. Их булава образована рыхло расположенными члениками, ,1 форма которых характерна для четковидных антенн.
Коленчатые антенны характеризуются вытянутым основным члеником, несущим расположенный под углом к нему относительно мелкочленистый жгутик.
Гребенчато-коленчатые усики сочетают в своем строении черты морфологии коленчатых и пластинчато-булавовидных антенн: характерную для первых основную часть и типичную для вторых пластинчатую булаву.
Неправильные антенны имеют в своем составе членики асимметричной (неправильной) формы.
Щетинконосные антенны, как правило, состоят из трех крупных члеников, последний из которых имеет характерную чувствительную щетинку, гладкую или перистую.
Основные типы грудных конечностей насекомых
Грудной отдел тела насекомых несет три пары конечностей. Исходными считаются конечности ходильного типа. При освоении насекомыми разных жизненных сред, иных способов передвижения либо при приобретении конечностями других функций изменилось и строение последних, носящее адаптивный характер. В составе конечности выделяются следующие части: тазик, вертлуг (трохантер), бедро, голень и многочлениковая лапка (рис. 113). Членистостьлапки является вторичной: ее членики не имеют самостоятельной мускулатуры, общий мышечный пучок пронизывает их, как нитка в бусах или четках. С особенностями строения конечностей основных типов следует ознакомиться, используя постоянные микроскопические препараты. Помимо описанных ниже, имеются и другие типы конечностей, менее распространенные, например присасывательные или прядильные.
Строение бегательных конечностей черного таракана (Blatta orien-talis). Бегательные конечности рассматриваются как модификация ходильных либо выделяются в отдельный самостоятельный тип. Бе-гательная конечность черного таракана состоит из тазика, вертлуга, бедра, голени и лапки (рис. 113, А). Тазик представляет собой основной членик, который неподвижно причленяется к грудному стерниту. Вертлуг у таракана, как и у многих других насекомых, миниатюрен, несколько смещен в сторону и располагается вдоль сочленения тазика и бедра. Бедро — наиболее крупный продолговатый членик. К нему причленяется также продолговатая, но более тонкая голень. Членистая лапка, сравнимая по длине с голенью, на вершине несет слегка
222
ImArthropoda
Класс Insecta 223

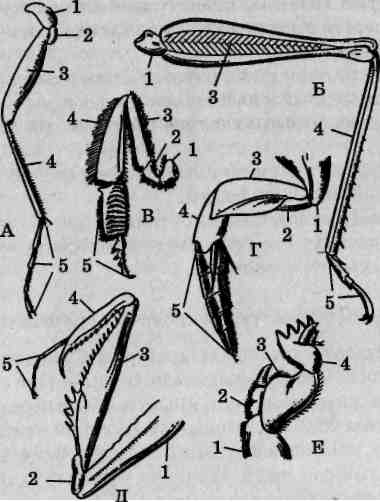
Рис. 113. Типы конечностей насекомых: А — ходильная; Б—прыгатель-ная; В — собирательная; Г— плавательная; Д— хватательная; Е — копательная; 1 — тазик; 2 — вертлуг; 3 — бедро; 4 — голень; 5—лапка
изогнутые коготки. Бегательные конечности обеспечивают быстрое перемещение по субстрату и удерживают тело высоко над его поверхностью, что необходимо, например, при передвижении по раскаленному солнцем песку. От ходильных конечностей бегательные отличаются относительно большей длиной и стройностью, а также строением лапки. Для конечностей ходильного типа характерны лапки с уплощенной нижней поверхностью, несущей большое количество волосков или щетинок и называемой подошвой.
Строение прыгательных конечностей кузнечиков (Tettigonia spp.). Прыгательные конечности обеспечивают передвижение прыжками, когда значительное расстояние преодолевается практически мгновенно. У кузнечиков и других прямокрылых прыгательными являются конечности третьей пары, у которых бедренно-голенный
сустав образует мощный рычаг. Бедра этих конечностей длинные и толстые (рис. 113, Б), с развитой мускулатурой. Голени также удлинены пропорционально длине бедер. Сходное строение имеет третья пара конечностей блох (Aphanipterd). У цикадовых (Cicadinea) «рабочим» является тазобедренный сустав.
Строение плавательных конечностей жуков-плавунцов (Dytiscidae). Плавательная (гребная) конечность характеризуется ограниченностью свободы движений большинства ее члеников относительно друг друга. В результате формируется единая гребная поверхность, работающая как весло. Следует обратить внимание на размеры и форму лапки (рис. 113, Е). У плавунцов она явственно уплощена и снабжена многочисленными длинными волосками. При выносе конечности вперед они прижимаются к поверхности голени, а при рабочем ходе расправляются и таким образом увеличивают гребную поверхность.
Плавательными могут быть все три пары конечностей (многие водные личинки жуков), средняя и задняя пары (имаго плавунцов) или только задняя (клопы-гребляки сем. Corixidae).
Строение хватательных конечностей водяного скорпиона (Nepa cinered). Хватательными у этого клопа являются передние конечности. По принципу работы они напоминают складной (перочинный) нож или капкан: удлиненные бедро и голень (рис. 113,5) при сведении зажимают добычу. Лапка, которая могла бы затруднять функционирование такой конечности, редуцирована, сохранив один членик. Поскольку хватательные конечности используются хищниками для захвата добычи, их иногда называют хищными. У обыкновенного богомола хватательные конечности устроены аналогично, но голень и бедро вооружены мощными заостренными хитиновыми зубцами, которыми насекомое способно пробить даже кожу человеческой руки (особенно, детской).
Строение копательных конечностей медведок (Gryllotalpa spp.). Копательными у медведок являются передние конечности. Они предназначены для рытья ходов в почве, зачастую пронизанной многочисленными мелкими корнями растений. Все элементы такой конечности сильно хитинизйрованы, укорочены и уплощены (рис. 113, Ж). Преимущественное развитие получает голень, которая снабжена мощными заостренными зубцами. Лапка подвергается редукции, но сохраняется.
Копательными конечностями обладают также жуки-навозники, некоторые клопы, жуки-усачи, короеды, то есть насекомые, которые по роду деятельности должны разгребать субстрат либо проделывать в нем ходы.
224
Тип Arthropoda
Класс Insecta
225
 Строение
собирательных конечностей медоносной
пчелы (Apis
Строение
собирательных конечностей медоносной
пчелы (Apis
mellifera). Собирательными у пчел являются задние конечности. Характерно строение голени и первого членика лапки (рис. \\Ъ,Д): внешняя поверхность голени гладкая, но по краям ее расположены многочисленные длинные изогнутые волоски, ограничивающие так называемую корзинку, которая служит для накопления и переноса пыльцы. Внутренняя поверхность очень крупного первого членика лапки усажена многочисленными волосками, за что это образование получило название щеточки. Пчела не занимается непосредственным сбором пыльцы из пыльников. Посещая цветки, она накапливает на опушенной поверхности тела пыльцевые зерна, а затем, совершая характерные движения, вычесывает пыльцу с помощью щеточки и помещает ее в корзиночку в виде так называемого пыльцевого комка, или обножки. Благодаря яркой окраске пыльцевые комки хорошо заметны у посещающих цветки пчел и шмелей.
Строение прицепных конечностей человеческой вши (Pediculus humanus). Прицепные конечности характерны для обитателей волосяного покрова млекопитающих и перьевого — птиц. Для этих эктопаразитов жизненно важно удержаться на хозяине, так как падение с него в большинстве случаев равнозначно гибели. У человеческой вши голень имеет специальный отросток, к которому вплотную подходит коготок лапки (рис. 113, Г). Сама лапка одночлениковая, сравнимый с нею по размерам коготок причленяется под углом, а просвет между голенью и лапкой соответствует толщине волоса. В результате формируется приспособление, напоминающее карабин в альпинистском снаряжении. Захватываемый волос ущемляется, чем обеспечивается надежная фиксация на нем эктопаразита. Захватывая волос за волосом, насекомое медленно перемещается в волосяном покрове хозяина.
Основные типы крыльев насекомых, жилкование крыла
Крылья насекомых по своему происхождению являются нечленистыми придатками второго и третьего грудных сегментов, образованными складками стенки тела. Форма, размеры и другие морфологические характеристики крыльев у представителей разных отрядов насекомых могут сильно различаться (рис. 114). Кроме основной, локомоторной функции, крылья могут выполнять и другие, что сопровождается соответствующими изменениями в их строении.
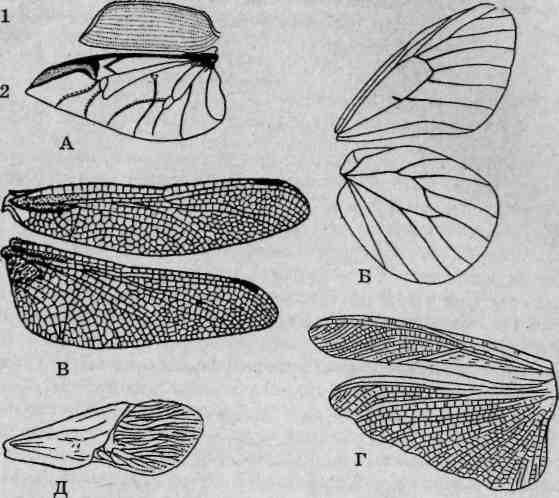
Рис. 114. Типы крыльев насекомых: А — жуков: 1 — жесткие (элитры); 2 — перепончатые; Б — чешуекрылых: чешуйчатые; В — стрекоз: сетчатые; Г— прямокрылых: кожистые (тегмины); Д— полужесткокрылых: полужесткие (гемиэлитры)
Наиболее распространенной является треугольная форма крыла. Различают основание, которым оно причленяется к грудному сегменту, противоположный ему апикальный (вершинный) угол и лежащий сзади нижний, или задний угол (рис. 115). Стороны крылового треугольника называются краями: костальным, или передним (соединяет основание с апикальным углом), терминальным, или наружным (апикальный и нижний углы), дорсальным, или задним (основание и нижний угол).
Если крылья прозрачны, не составляет труда рассмотреть их жилкование (расположение трахеи). Жилки представляют собой разветвленные трахеи. Если среди них невозможно выделить крупнейшие основные и жилкование выглядит как система мелких ячеек, такие крылья называются сетчатыми. Они характерны для стрекоз, поденок
226 Тип Arthropods
Класс Insecta 227
3
Рис.
115. Общая
схема строения крыльев насекомых:
1
—
основание крыла; 2
—
задний
угол; 3—
вершина;
4—
передний
» край;
5
—
наружный край; 6
—
внут-
ренний
край крыла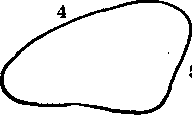
стальные). Далее следует ветвящаяся радиальная, а за нею — также ветвящаяся медиальная. Следующая, кубитальная, жилка разветвляется надвое. Последующие анальные жилки не ветвятся. Югаль-ная область крыла, наличие которой обеспечивает возможность насекомому складывать крылья вдоль тела, включает целый набор не-ветвящихся югальныхжилок. Для большинства насекомых характерны более или менее существенные отклонения от описанного
и сетчатокрылых. У большинства других насекомых на прозрачных крыльях нетрудно выделить ограниченный круг радиально расходящихся от основания основных жилок. Эти крылья носят название перепончатых. Таковыми являются обе пары крыльев насекомых отряда перепончатокрылых, передняя пара — двукрылых, задняя — жесткокрылых и полужесткокрылых. У ручейников крылья несут множество волосков, за что отряд и получил свое название — волосокрылые. Крылья бабочек (чешуекрылых) носят название чешуйчатых, так как их поверхность покрыта слоем черепицеобразно расположенных чешуек. Жилкование таких крыльев становится явным после их смачивания авиационным бензином или аналогичным растворителем. Восполь-. зовавшись бинокуляром, следует рассмотреть поверхность крыльев ручейников и бабочек. Передние крылья тараканов и прямокрылых носят название кожистых надкрылий, или тегмин, что вполне соответствует их механическим свойствам. У жуков передняя пара крыльев интенсивно склеротизирована до рогоподобного состояния. Такие крылья называются жесткими, или элитрами. Основная функция тегмин и элитр — защитная. У клопов склеротизировано лишь основание переднего крыла, тогда как вершина его остается перепончатой и называется мембраной. Она может сложным образом складываться, оставаясь, наряду с задними крыльями, защищенной более жесткими основаниями. Такие крылья получили название гемиэлитр (полужестких). Отсюда и второе название отряда клопов — полужесткокрылые. У двукрылых вторая пара крыльев модифицирована в лопаткообразные жужжальца. Они выполняют функцию стабилизаторов полета и активизируют работу крыловых мышц передней пары крыльев.
По характеру жилкования крыла часто определяют таксономическую принадлежность насекомых. Во избежание путаницы зоологами приняты стандартные схемы обозначения отдельных жилок и образуемых ими ячеек. В русскоязычной специальной литературе используется следующая генерализированная схема жилкования (рис. 116): ближе всего к переднему (костальному) краю крыла расположена костальная жилка. За ней следует субкостальная, которая у своей вершины подразделяется на две (первую и вторую субко-
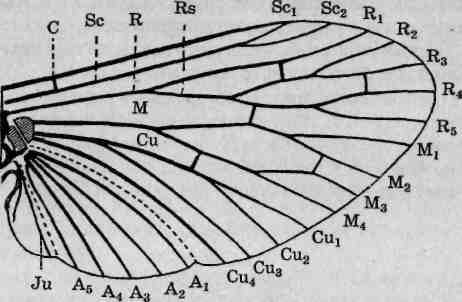
Рис. 116. Генерализированная схема жилкования крыльев насекомых: С — костальная жилка; Scj—Sc2 — субкостальные жилки; R; — Rs— радиальные жилки; Л/, — М4 — медиальные жилки; Си} — Cu4 — кубитальные жилки; А, — А5 — анальные жилки; Ju — югальная жилка
варианта жилкования. Как пример можно рассмотреть особенности жилкования крыла боярышницы. Для знакомства с разнообразием жилкования используются препараты крыльев насекомых разных отрядов.
Основные функционально-морфологические типы ротовых аппаратов насекомых
Ротовые аппараты насекомых весьма разнообразны по своему строению, что определяется в первую очередь характером пищи и способом ее потребления. Насекомые используют в пищу самые различные пищевые субстраты, и по этой причине ротовой аппарат претерпевает соответствующие морфологические адаптивные изменения. Основными функционально-морфологическими типами ротовых аппаратов
228
Тип Arthropoda
Класс Insecta
229
 насекомых
считаются: грызущий, лакающий
(грызуще-лижущий), сосущий
лепидоптероидный, лижущий мускоидный,
колюще-сосущие ге-миптероидный
и диптероидный.
насекомых
считаются: грызущий, лакающий
(грызуще-лижущий), сосущий
лепидоптероидный, лижущий мускоидный,
колюще-сосущие ге-миптероидный
и диптероидный.
Строение грызущего ротового аппарата черного таракана (Blatta orientate). Этот тип ротового аппарата характерен для наиболее древних насекомых и считается исходным, тогда как прочие возникли в результате его модификации. Ротовой аппарат черного таракана отличается относительно невысокой степенью склеротизации, в связи с чем эти насекомые могут измельчать лишь сравнительно мягкие пищевые субстраты. Твердую пищу они предварительно должны смачивать слюной и только после этого могут ее измельчить и съесть.
Сверху ротовые части прикрывает непарная верхняя губа (лабрум), представляющая собой складку стенки тела в виде широкой пластинки с прямоугольным основанием и закругленной вершиной (рис. 117). Она подвижно причленена к головной капсуле под наличником. Верхняя губа выполняет роль передней стенки предротовой полости.
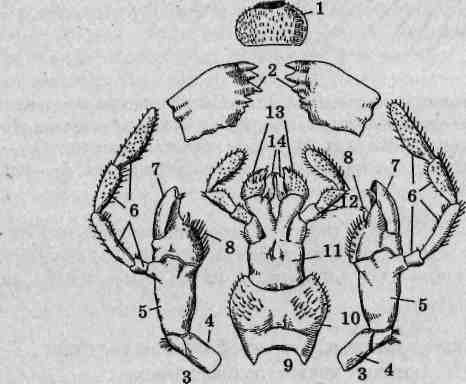
Рис. 117. Ротовой аппарат черного таракана (Blatta orientalis): 1 — верхняя губа; 2 — жвалы (верхние челюсти); 3 — нижние челюсти; 4— основной членик; 5— стволик; 6— челюстной щупик; 7— наружная жевательная лопасть; 8 — внутренняя жевательная лопасть; 9— нижняя губа; 10— подподбородок; 11 — подбородок; 12— нижнегубной щупик; 13— придаточные язычки; /'/—язычки
Верхние челюсти (мандибулы, жвалы) представляют собой пару коротких лопастевидных нечленистых пластинок. Они имеют темную окраску и интенсивно склеротизированы. Внутренние края жвал снабжены несколькими крупными зубцами. Мандибулы не являются строго симметричными, поскольку зубцы одной из них должны совпадать с промежутками между зубцами другой.
Нижние челюсти (максиллы-I) сохранили парность и членис-тость строения. С головной капсулой они соединены основным члеником, или подвеском (кардо), к которому причленяется более массивный стволик (стипес). У вершины стволика с внешней стороны на бугорке сидит четырехчлениковый нижнечелюстной (максил-лярный) щупик, выполняющий чувствительную функцию. Широкая вершина стволика несет также пару жевательных лопастей. Внутренняя жевательняя лопасть (лациния) сильно склеротизирована, снабжена хитиновыми зубцами и щетинками, она принимает непосредственное участие в измельчении пищи. Наружная жевательная лопасть (галеа) слабо склеротизирована и покрыта многочисленными волосками.
Нижняя губа (лабиум) непарная, но образована в результате слияния основных члеников максилл-П (максиллул) и сохраняет членистое строение. В частности, лежащий в ее основании широкий сердцевидный подподбородок (субментум) соответствует подвескам нижних челюстей. К нему причленяется более узкий подбородок (мен-тум), образованный в результате слияния стволиков. Подбородок несет щупики и язычки. Собственно язычки (глоссы), расположенные по центру, соответствуют внутренним жевательным лопастям, а придаточные язычки (параглоссы), занимающие промежуточное положение между язычками и нижнегубными щупиками, — наружным жевательным лопастям максилл-1. Нижнегубные (лабиальные) щупики короче нижнечелюстных, состоят из трех члеников и выполняют чувствительную функцию, причем более специализированы к хеморецепции. Имеющая плоскую форму нижняя губа замыкает снизу предротовую полость и служит для слизывания жидкой пищи. На ее внутренней стороне имеется небольшой подвижный подглоточ-ник (гипофарингс), функционально аналогичный языку большинства млекопитающих. По своему происхождению он представляет непарный вырост стенки тела. Ввиду малых размеров и отсутствия склеротизации гипофарингс обычно не принимается во внимание при изготовлении демонстрационных микроскопических препаратов.
230
Тип Arthropoda
Класс Insecta 231
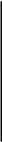
 Строение
лакающего (грызуще-лижущего) ротового
аппарата шмеля (Bombus
sp.). Такой
ротовой аппарат характерен для пчелиных,
имаго которых
собирают нектар из цветков, и по строению
(рис. 118) напоминает рассмотренный
выше грызущий, где все элементы в той
или иной
степени подверглись модификации.
Строение
лакающего (грызуще-лижущего) ротового
аппарата шмеля (Bombus
sp.). Такой
ротовой аппарат характерен для пчелиных,
имаго которых
собирают нектар из цветков, и по строению
(рис. 118) напоминает рассмотренный
выше грызущий, где все элементы в той
или иной
степени подверглись модификации.
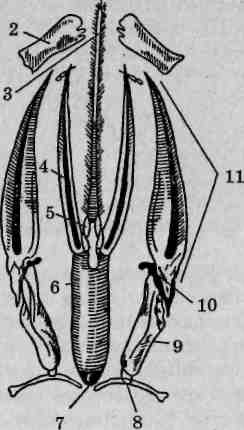
Рис. 118. Ротовой аппарат шмеля (Bombus): 1 — верхняя губа; 2 — верхняя челюсть; 3 — язычок; 4 — нижнегубной щупик; 5 — наружная лопасть нижней губы; 6— подбородок; 7 — под-подбородок; 8 — основной членик; 9 — стволик; 10 — нижнечелюстной щупик; 11 — нижняя челюсть
Верхняя губа представляет собой короткую, но широкую хитини-зированную пластинку, верхний закругленный край которой покрыт многочисленными волосками.
Верхние челюсти внешне напоминают таковые черного таракана, но отличаются продолговатой формой. Они не играют существенной роли в процессе потребления пищи, но активно используются для обработки воска, для перемещения или разгрызания травинок и других предметов при строительстве гнезда. Известна способность шмелей и пчел делать боковые надкусы в длинных венчиках цветков, имеющих труднодоступные удлиненные нектарники. У медоносной пчелы мандибулы имеют характерный желобок и используются для сбора пыльцы.
Нижние челюсти и нижняя губа объединены в хоботок. Основания максилл имеют палочковидную форму. К ним причленяются хорошо развитые стволики. Наружные жевательные лопасти хорошо развиты, имеют форму вытянутых клиновидных пластинок, способных при необходимости желобоподобно изгибаться. Внутренние жевательные лопасти, как и нижнечелюстные шупики, рудиментарны и располагаются у оснований наружных жевательных лопастей.
Подподбородок нижней губы имеет форму треугольника со слегка вогнутыми сторонами и причленяется к головной капсуле. Подбородок удлиненный, бочонкообразный. К нему причленен единый язычок (глосса), имеющий ребристую поверхность и снабженный многочисленными волосками. В толще язычка проходит канал, служащий для всасывания жидкой пищи. Придаточные язычки имеют вид коротких желобчатых пластинок. Сбоку от них отходят нижнегуб-ные щупики, которые претерпели морфологическую модификацию и сменили основную функцию. Первый членик щупика развит лучше других, по длине сравним с наружными жевательными лопастями нижних челюстей, уплощен и имеет форму мелкого желоба. Остальные три членика щупика невелики по размерам и сохранили стандартное строение.
Хоботок является весьма эффективным приспособлением для потребления нектара. Смыкающиеся наружные жевательные лопасти нижних челюстей образуют дорсальную и боковые стенки, а первые членики нижнегубных щупиков — вентральную стенку трубчатого канала хоботка. Внутри размещается язычок, который при потреблении животным жидкой пищи вибрирует. В этом отношении он напоминает ершик для мытья узкой посуды. Мощная глоточная мускулатура создает разрежение, что в комплексе и обеспечивает поступление пищи. Малые количества нектара, находящиеся к тому же в наиболее труднодоступных местах, удается достать с помощью длинного язычка и откачать по его внутреннему каналу.
Строение гемиптероидного колюще-сосущего ротового аппарата клопов (Hemipterd). Данный тип ротовых аппаратов (рис. 119) характерен для всех представителей отряда полужесткокрылых, или клопов (Hemipterd). Он специализирован для потребления жидкой пищи, находящейся под покровными тканями животных или растений.
Верхняя губа имеет вид небольшой, заостренной на вершине пластинки, причлененной к наличнику.
Верхние и нижние челюсти преобразованы в нечленистые хитиновые стилеты. В совокупности мандибулы и максиллы
232
Тип Arthropoda
Класс Insecta
233

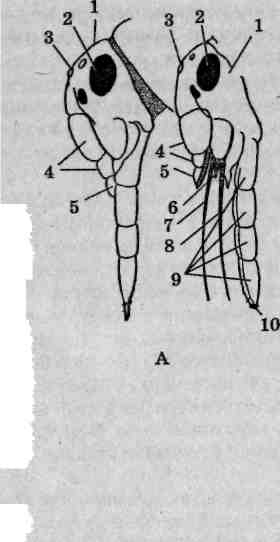
Рис. 119. Ротовой аппарат клопов (Hemiptera): А — общая схема строе- I ния, вид сбоку: 1 — головная капсула; 2— фасеточный глаз; 3—| лоб; 4 — двураздельный наличник; 5 — верхняя губа; б — " подглоточник; 7 — мандибулы; 8 — максиллы; 9-10 — нижняя губа (9— членики нижней губы, 10— желоб); Б — стадии проникновения стилетов в ткани хозяина: 1 — сложенные вместе челюсти, прокалывающие ткань; 2 — нижняя губа; В — схема строения хоботка на поперечном срезе: / — мандибулы; . 2 — максиллы; 3 — дорсальный (пищевой) канал; 4 — вентральный (слюнной) канал
образуют подобие наборной иглы, пронизанной несколькими внутренними каналами. Мандибулы, имеющие на срезе форму полуколец, составляют обкладку иглы. Концы их заострены, а в ряде случаев несут также дополнительные зубцы и предназначены для прокалывания покровов. Максилллы в соприкосновении друг с другом образуют пару крупных каналов (рис. 119, В), обеспечивающих эф-
фективную транспортировку жидкости. По вентральному каналу в рану подается слюна, содержащая пищеварительные ферменты. Дорсальный канал предназначен для всасывания жидкой пищи.
Нижняя губа выполняет функцию кожистого чехла (ножен) для стилетоподобных мандибул и максилл. Она представляет собой продолговатый отросток, образованный тремя—четырьмя члениками и снабженный спереди желобком для стилетов, которые могут свободно скользить по нему. В состоянии покоя стилеты полностью скрыты в канале нижней губы (рис. 119, Б). При погружении их в ткани хозяина нижнегубной чехол образует направленный назад колено-образный или иной формы изгиб. У некоторых равнокрылых стилеты длиннее не только нижней губы, но и всего тела своего владельца и в покое свернуты широкой петлей или спиралью внутри головной капсулы. Опистогнатический тип расположения ротового аппарата позволяет этим насекомым развивать на вершине аппарата проникновения давление, достаточное для преодоления весьма прочных покровов их жертв.
Строение диптероидного колюще-сосущего ротового аппарата кровососущих комаров (сем. Culicidae). Принадлежащие к отряду двукрылых (Diptera) комары указанного семейства располагают колюще-сосущим ротовым аппаратом несколько иного строения. В отличие от гемиптероидного типа, в его составе присутствует шесть стилетооб-разных образований (рис. 120).
Верхняя губа имеет вид длинной тонкой трубки с косо срезанным, заостренным концом. Стенки этой трубки не сплошные: вдоль внутренней (вентральной) ее стороны идет шов с неспаянными краями. Кроме того, толщина стенок крайне неравномерна: спереди (с дорсальной стороны) она больше, чем сзади (с вентральной стороны), вокруг шва. К последнему прилегает комплекс сложенных определенным образом остальных стилетоподобных элементов, имеющих на срезе слегка вогнутую форму. Мандибулы превращены в тонкие жесткие хитиновые щетинки, заостренные на вершине и зазубренные по краям. Нижние челюсти превратились в примерно такие же щетинки, но сохранили длинные и тонкие членистые щупики. Гипофарингс также трансформировался в желобоподобную, стилетообразную щетинку. Внутри он пронизан тонким каналом, по которому в рану подается слюна насекомого.
Весь описанный комплекс элементов комбинируется в стержнепо-добное образование, которое в нерабочем состоянии спрятано в чехле, образованном нижней губой. В отличие от клопов, у комаров она нечлениста и содержит больше мягких тканей. В результате можно
234
Тип Arthropoda
Класс Insecta
235

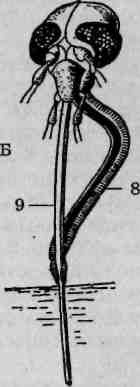
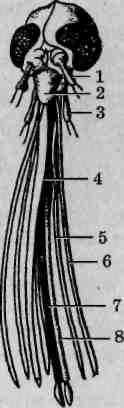
Рис. 120. Ротовой аппарат настоящего комара (Culex): A — общая схема строения; Б — проникновение стилета в ткани хозяина; 1 — основания антенны; 2—лобный щиток; 3— основание челюстного щупика; 4 — верхняя губа; 5 — максиллы; 6 — мандибулы; 7 — подглоточник; 8 — нижняя губа; 9 — сложенные и вводимые вместе верхняя губа, мандибулы, максиллы, подглоточник
заключить, что гемиптероидныи и диптероидныи варианты ротовых аппаратов колюще-сосущего типа устроены и функционируют сходным образом.
Строение сосущего ротового аппарата чешуекрылых (Lepidopterd) (рис. 121). Этот тип ротового аппарата весьма характерен для представителей отряда чешуекрылых, или бабочек. Специализация к потреблению жидкой пищи (нектара, вытекающего из ран деревьев сока и пр.) привела к значительным изменениям по сравнению с исходным вариантом строения.
Верхняя губа имеет вид небольшой пластиночки или складки, причлененной к наличнику, и в функционировании ротового аппарата участия не принимает. Верхние челюсти у большинства представителей отряда утрачены.
Нижние челюсти в наименьшей степени подверглись редукции. Хорошо развиты наружные жевательные лопасти в виде длинных,
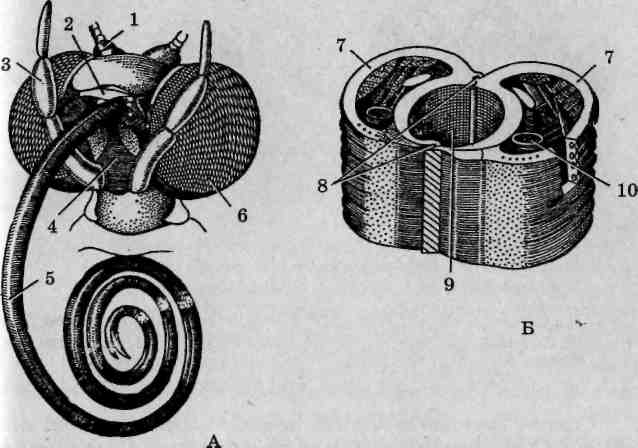
Рис. 121. Ротовой аппарат чешуекрылых (Lepidopterd): А — голова с расправленным хоботком; Б — участок хоботка на срезе; 1 — основание антенны; 2 — верхняя губа; 3 — нижнегубной щупик; 4 — нижняя губа; 5— хоботок; 6— фасеточный глаз; 7— правая и левая нижние челюсти; 8 — места соединения челюстей; 9 — полость хоботка; 10— трахея
желобообразных пластинок. Вместе они образуют трубку — хоботок, через канал которого всасывается жидкая пища. В силу своей эластичности хоботок свертывается в почти плоскую спираль типа часовой пружины и в покое подогнут под грудь. Для развертывания хоботка в него под давлением нагнетается гемолимфа, в этом процессе участвует и внутренняя мускулатура.
Нижняя губа представляет собой небольшую пластинку, расположенную позади нижних челюстей. Хорошо развиты нижнегубные щупики, имеющие типичное строение и выполняющие чувствительную функцию.
Таким образом, специализация к питанию свободной от защитных оболочек жидкой пищей привела у чешуекрылых к преобразованию почти всех элементов ротового аппарата.
236
Тип Arthropoda [
Класс Arachnida
237
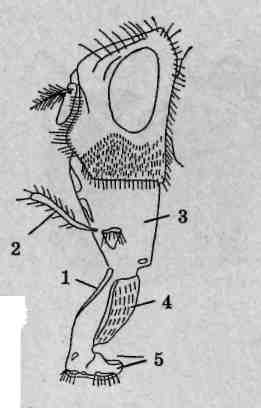 Рис.
122. Ротовой
аппарат комнатной мухи (Musca
domestica): 1
—
верхняя губа; 5 2
—
максиллярный щупик; 3
—
рост-, рум;
4—
гаустеллум;
5—
лабеллумы
Рис.
122. Ротовой
аппарат комнатной мухи (Musca
domestica): 1
—
верхняя губа; 5 2
—
максиллярный щупик; 3
—
рост-, рум;
4—
гаустеллум;
5—
лабеллумы
Строение лижущего мускоидного ротового аппарата (хоботка) высших мух. Среди рассматриваемых типов он наиболее далек от исход-§ ного варианта строения (рис. 122).
Основание хоботка называется рострумом. В его образовании, Г помимо разросшихся краев головной капсулы, принимает участие -наличник. Спереди он несет нижнечелюстные щупики. Рострум переходит в гаустеллум, образованный подглоточником (гипофа- j рингсом). Спереди он защищен приросшей к его поверхности верхней I губой. Далее следует парный или двухлопастный лабеллум, в форми- | ровании которого основная роль принадлежит нижнегубным щупикам. . Рострум, гаустеллум и лабеллумы сочленены подвижно, в состоянии I покоя они складываются двумя коленами под головной капсулой и малозаметны со стороны.
В рабочем состоянии лабеллумы имеют вид полукруглых присо-; сок с расположенным между ними центральным отверстием, ведущим в глотку. К этому отверстию и идущему от него к глотке каналу подходят мелкие канальцы, называемые псевдотрахеями. На другом:. конце они открываются микропорами на вентральной поверхности ла-беллума. Края центрального отверстия имеют хитиновые зубцы («зу-I бы»), способные дробить твердые субстраты (рис. 123). Питающаяся на поверхности твердого субстрата муха сначала смачивает его слю- ; ной. Если он оказывается нерастворимым, она пытается дробить его .
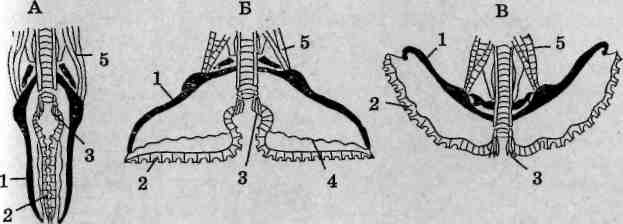
Рис. 123. Разворачивающиеся лабеллумы ротового аппарата высших двукрылых (Diptera): А —в состоянии покоя; Б— в процессе фильтрации жидкой пищи; В — в процессе обскребывания субстрата; / — склериты; 2 — псевдотрахеи; 3 — «зубы»; 4 — гибкая мембрана; 5 — мышцы
поверхность. Когда насекомое втягивает полученный раствор или жидкую кашицу, лабеллумы функционируют наподобие поролоновой губки. Затем животное повторяет цикл на новом месте. У осенней жигалки (Stomoxys calcitrans), мух-цеце (Glossina) вооружение краев центрального отверстия представлено в виде плоских пружинящих лезвий, с помощью которых животные способны без труда вырезать характерную лунку даже в очень плотных кожных покровах, например крупного рогатого скота.
Описанными типами ротовых аппаратов не исчерпывается их разнообразие у насекомых. Общим является присутствие в их составе соответствующим образом модифицированных конечностей и вторичных придатков головного отдела тела.
Подтип Chelicerata — хелицеровые
Класс Arachnida — паукообразные
Класс объединяет наземных и вторичноводных членистоногих. Первая пара головных конечностей — исходно клешневидные хел и -Церы, занимающие предротовое местоположение. Тело подразделяется на два отдела: просому и опистосому, последняя объединяет в своем составе мезосому и метасому (рис. 124). Просома несет шесть пар конечностей, включая хелицеры. Органы дыхания (если имеются)
238
Тип Arthropods
Класс Arachnida
239

А Б В Г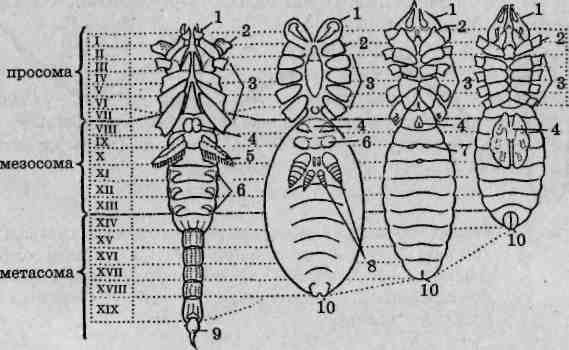
Рис. 124. Строение арахнид (вид с брюшной стороны): А — скорпиона; Б — паука; В — сольпуги; Г— примитивного акариформного клеща; I—XIX — сегменты тела; 1 — хелицеры; 2— педипалыгы; 3— ходильные конечности; 4— половое отверстие; 3 — гребневидные органы (половые гребешки); 6 — стигмы легких; 7 — стигмы трахей; 8— паутинные бородавки; 9— хвостовой придаток; 10 — анальное отверстие
воздушного типа. Органами выделения служат мальпигиевы сосуды. На лабораторных занятиях целесообразно остановиться на рассмотрении особенностей внешней морфологии представителей пяти подклассов паукообразных: скорпионов, сольпуг, пауков, ложноскорпионов и клещей. С особенностями строения сенокосцев, таксономическим разнообразием пауков, типами повреждений растений клещами-фитофагами удобнее ознакомиться в ходе летней учебной практики.
Подкласс Scorpiones — скорпионы
Представители подкласса — крупные хелицеровые со слитной го ловогрудью (цефалотораксом) и членистым брюшком (абдоменом), состоящим из передне- и заднебрюшия. Тельсон снабжен ядовитой железой. Хелицеры и педипальпы клешневидные. Дыхание легочное. Представители рассматриваемого подкласса раздельнополы, развитие у них прямое. Скорпионы теплолюбивы, в пределах Восточной Европы
встречаются в Прикаспии, Предкавказье, Причерноморье и Средиземноморье. Хищники.
Buthus eupeus — пестрый скорпион
Материал. Пестрый скорпион, как и другие представители семейства Buthidae, распространен в Нижнем Поволжье, Средней Азии и Закавказье, где местами многочислен. Для Причерноморья характерны представители другого семейства скорпионов — Chactidae: в Крыму это эндемичный Euscorpius tauricus, на Черноморском побережье Кавказа — Euscorpius italicus, в Западном Закавказье — Euscorpius mingrelicus. Светлое время суток эти животные проводят в укрытиях. Их можно отловить под камнями, отслаивающейся корой деревьев, в трещинах скал и стен домов. Для сбора скорпионов используют длинный пинцет. Из-за агрессивности и склонности к каннибализму их нельзя держать по нескольку экземпляров в одном сосуде. Укус скорпиона не представляет прямой опасности для взрослого человека, однако во избежание анафилактического шока и других клинических реакций следует проявлять максимальную осторожность. Собранных животных немедленно помещают в заправленную морилку. Хранят материал в 70° этаноле.
Внешнее строение. Скорпионы принадлежат к числу одних из самых крупных (длина тела взрослых пестрых скорпионов достигает 65 мм) наземных членистоногих. Тело подразделено на головогрудь (цефалоторакс) и брюшко (абдомен). Граница между ними, если смотреть со спинной стороны (рис. 125), вполне явная. Со спинной стороны головогрудь прикрыта карапаксом, образуемым слившимися тергитами (спинными пластинками) шести головогрудных сегментов и покровами акрона. Воспользовавшись ручной лупой либо бино-куляром, можно рассмотреть располагающиеся в передней части головогруди простые глаза: по центру размещена пара медианных глаз, по переднему краю —фронтальных, по бокам —латеральных. На нижней стороне головогруди выделяется вентральный щит — стернум, образовавшийся в результате слияния брюшных пластинок (стернитов) двух последних грудных сегментов.
Брюшной отдел тела состоит из внешне хорошо отграниченных двенадцати настоящих сегментов и тельсона. Различают переднебрю-шие (мезосому), образованное семью сегментами, и заднебрюшие (метасому), в состав которого входят остальные пять сегментов и тельсон. Визуально тело скорпиона делится на широкий массивный корпус, включающий головогрудь и переднебрюшие, и узкое, длинное и гибкое
![]()
Ьъ_
ImArthropoda
a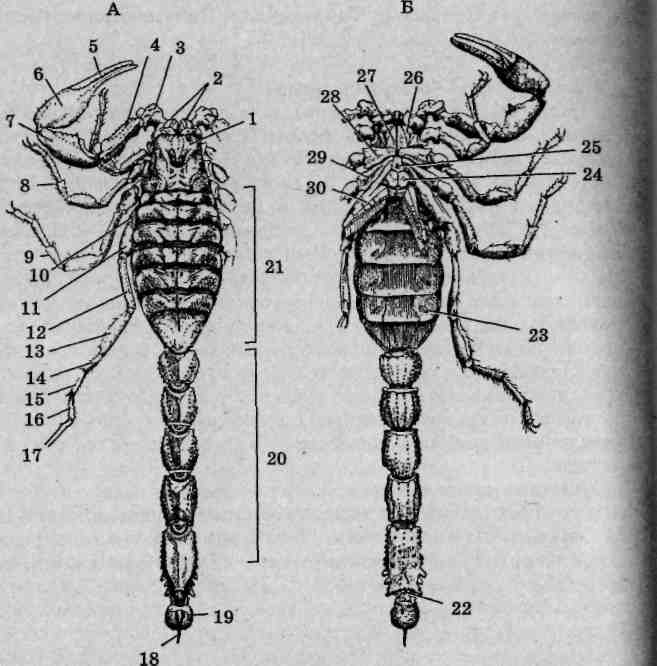
Рис. 125. Пестрый скорпион (Buthus eupeus): A — вид со спинной стороны; Б— вид с брюшной стороны; 1 — головогрудь; 2—хелицеры; 3 — вертлуг педипальпы; 4 — бедро педипальпы; 5— подвижный палец; 6 — ладонь; 7 — голень педипальпы; 8 — конечность II пары; 9— конечность III пары; 10-17— конечность IV пары (10— тазик; 11— вертлуг; 12— бедро; 13— голень; 74-основной членик лапки; 15— I члениклапки; 16— II членик лапки; 17— коготки); 18— ядовитая игла; 19— тельсон;20— заднебрюшие; 21 — переднебрюшие; 22 — анальное отверстие; 23 — стигма; 24 — половая крышечка; 25 — стернум; 26— челюстная лопасть ноги I пары; 27— челюстная лопасть конечности II пары; 28— тазик конечности II пары; 29— тазик конечности III пары; 30— гребневидный орган
Класс Arachnida
заднебрюшие, порой неправильно называемое «хвостовой час К последнему брюшному сегменту причленяется округлый тельсон несущий сверху изогнутую, заостренную на вершине хитиновую иглу — ядовитое жало скорпиона. Внутри тельсона находится парная ядо-в итая железа, для выведения секрета которой служит идущий внутри иглы канал. Скорпионы используют ядовитое жало как орудие защиты и нападения. Потревоженное животное изгибает брюшко, удерживая его конец над головогрудью — это типичная поза угрозы. Кроме зашиты от врагов, яд широко используется для быстрого умерщвления жертв: помимо различных членистоногих ими могут быть мелкие рептилии и даже детеныши мелких млекопитающих. Анальное отверстие расположено на брюшной стороне тела фактически на сочленении последнего сегмента заднебрюшия и тельсона. У некоторых экземпляров оно хорошо заметно благодаря окружающим его лопастевидным возвышениям, представляющим собой вывернутые наружу стенки прямой кишки.
Состав и особенности строения конечностей. Первой парой конечностей головогруди являются короткие клешневидные хелицеры. Расположены они фронтально (рис. 125, Б), оставляя позади ротовое отверстие. Хелицеры трехчленисты, предпоследний их членик несет специальный выступ, играющий роль неподвижного пальца клешни, как у речных раков. Функцию подвижного пальца выполняет последний членик хелицер. Основное назначение этих образований — механическая обработка пищевых объектов.
Вторая пара конечностей, педипальпы (ногочелюсти), выделяется среди прочих своей длиной. Они шестичлениковые, причем последние два членика образуют мощные клешни. Помимо обороны и, главное, захвата добычи, педипальпы выполняют осязательную функцию.
Последующие, третья - шестая, пары конечностей являются ходильными и на своих вершинах несут коготки. Они лишены каких-либо следов двуветвистости, задние развиты несколько лучше передних.
Тазики педипальп и всех ходильных конечностей вошли в состав брюшной стенки тела, утратив вследствие этого подвижность. Тазики двух первых пар ходильных конечностей снабжены специальными отростками — челюстными лопастями. Они легко различимы при осмотре брюшной поверхности головогруди: первая пара имеет вид треугольных, вторая — клиновидных пластинок. Лопасти окаймляют щелевидное ротовое отверстие, образуя в совокупности нижнюю губу. Скорпионы способны поглощать только жидкую и кашицеобразную пищу, для этого им необходимо предварительно ее измельчить и обработать слюной.
9Зак. 1602
242
Тип Arthropoda
Класс Arachnida
243
 Брюшной
отдел тела у скорпионов лишен конечностей
типичного
строения, но первые его сегменты несут
придатки, рассматриваемые
как модифицированные конечности. Первый
брюшной сегмент \
снабжен
округлыми образованиями, прикрывающими
расположенные <
здесь
половые отверстия и называемыми половыми
крышечками. У
хранившихся в этаноле экземпляров они
выделяются на общем фоне своей
светлой окраской. У некоторых самок
под половыми крышеч- | ками
могут быть обнаружены черные пластинки.
Это сперматоклей- ^* трумы,
которыми самцы запечатывают половые
отверстия после завершения
процесса спаривания. Они могут сохраняться
вплоть до времени
окончательного созревания яиц. На
втором брюшном сегменте
располагаются гребневидные придатки,
которые несут большое
число рецепторов и участвуют в ритуале
спаривания скорпионов.
На последующих четырех сегментах брюшка
по бокам имеются щелевидные
отверстия — стигмы, или дыхальца. Они
ведут в ле-i
точные
мешки, которые, как полагают специалисты,
по своему происхождению
представляют впятившиеся внутрь
дыхательные конечности.
У экземпляров, находившихся продолжительное
время в воде, края
стигмальных отверстий могут быть
заметно вздутыми. Послед-ним,
седьмым, сегментом переднебрюшия, как
и всеми сегментами заднебрюшия,
конечности полностью утрачены. Таким
образом, ко-J
нечности
брюшного отдела в большей степени,
нежели головогруд- [ ного,
подверглись трансформации либо редукции.
Брюшной
отдел тела у скорпионов лишен конечностей
типичного
строения, но первые его сегменты несут
придатки, рассматриваемые
как модифицированные конечности. Первый
брюшной сегмент \
снабжен
округлыми образованиями, прикрывающими
расположенные <
здесь
половые отверстия и называемыми половыми
крышечками. У
хранившихся в этаноле экземпляров они
выделяются на общем фоне своей
светлой окраской. У некоторых самок
под половыми крышеч- | ками
могут быть обнаружены черные пластинки.
Это сперматоклей- ^* трумы,
которыми самцы запечатывают половые
отверстия после завершения
процесса спаривания. Они могут сохраняться
вплоть до времени
окончательного созревания яиц. На
втором брюшном сегменте
располагаются гребневидные придатки,
которые несут большое
число рецепторов и участвуют в ритуале
спаривания скорпионов.
На последующих четырех сегментах брюшка
по бокам имеются щелевидные
отверстия — стигмы, или дыхальца. Они
ведут в ле-i
точные
мешки, которые, как полагают специалисты,
по своему происхождению
представляют впятившиеся внутрь
дыхательные конечности.
У экземпляров, находившихся продолжительное
время в воде, края
стигмальных отверстий могут быть
заметно вздутыми. Послед-ним,
седьмым, сегментом переднебрюшия, как
и всеми сегментами заднебрюшия,
конечности полностью утрачены. Таким
образом, ко-J
нечности
брюшного отдела в большей степени,
нежели головогруд- [ ного,
подверглись трансформации либо редукции.
Подкласс Solpugida (=SoliJugae) — сольпуги
Подкласс объединяет преимущественно крупных арахнид, внеш- ' ним обликом напоминающих пауков. Степень расчлененности тела взрослых особей сольпуг выше, чем у всех остальных групп пауко-: образных. Хелицеры клешневидные. Педипальпы ходильно-осязательного типа. Дыхание трахейное. Все сольпуги — хищники.
Для ознакомления с особенностями сегментации тела представителей отряда можно использовать смонтированный сухой материал либо влажные препараты взрослых особей любого вида фаланг и других сольпуг.
Galeodes araneoides — обыкновенная фаланга
Материал. Обыкновенная фаланга характерна для степей и предго-' рий Крыма, Приазовья, Предкавказья и Нижнего Поволжья. На большей; части территории Средней Азии вполне обычна, а местами массовы
видом является закаспийская сольпуга (Galeodes caspius). Представители рода Galeodes активны в темное время суток. Их привлекает свет, чем пользуются для сбора этих животных. Мелкая светлоокрашенная солнцелюбивая сольпуга (Paragaleodes heliophilus) принадлежит к числу дневных форм. В среднеазиатских степях и полупустынях этих паукообразных можно наблюдать перемещающимися по поверхности почвы даже в знойный поддень. Для учебных целей пригодны все вышеупомянутые представители подкласса. Собранных сольпуг анестезируют эфирами, после чего требуется специальная обработка во избежание загнивания тканей мясистого брюшка. При хранении в жидких фиксаторах (80° этанол, 4% раствор формалина) обязательно с помощью шприца с тонкой иглой в брюшко делают несколько инъекций фиксатора. При монтировании сольпуг для сухих коллекций через небольшой разрез на нижней поверхности брюшко освобождают от мягких тканей и набивают ватными шариками, как это рекомендуется для крупных экземпляров пауков или жуков-маек.
Внешнее строение. Фаланги рода Galeodes принадлежат к числу наиболее крупных: длина тела взрослых особей Galeodes araneoides достигает 50 мм. Головогрудной отдел тела у сольпуг состоит из слитного пропельтидия (образован слиянием четырех передних сегментов) и двух свободных сегментов (рис. 126). Со спинной стороны пропельтидий покрыт мощным склеротизированным головным щитом. У его переднего края расположен глазной бугорок, несущий пару выпуклых простых глаз. Латеральные глаза у сольпуг подвержены атрофии.
Брюшко овально-цилиндрическое, состоит из десяти сегментов с хорошо различимыми границами. Туловище, а также придатки тела сольпуг покрыты густыми волосками и щетинками различной длины, формы и жесткости. На нижней стороне первого брюшного сегмента можно обнаружить половое отверстие, имеющее вид продольной щели, прикрытой боковыми створками.
Состав и особенности строения конечностей. Первая пара конечностей головогруди, хелицеры, получили у сольпуг преимущественное развитие. Размерами они сравнимы с пропельтидием и представляют собой направленные вперед, вертикально поставленные мощные клешневидные образования. Вершины их заострены, а внутренние поверхности несут многочисленные зубцы. Хелицеры являются мощным орудием защиты и нападения, ими сольпуги захватывают, умерщвляют и расчленяют добычу. Крупные взрослые особи могут даже прокусить кожу человека. Бытует мнение о ядовитости сольпуг, но оно ошибочно,
244
IvmArthropoda
Класс Arachnida
245

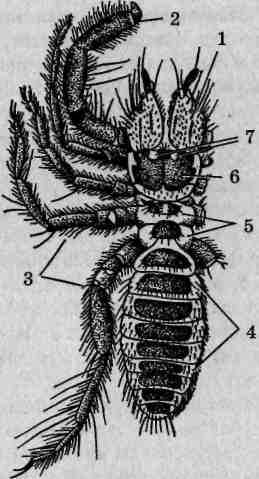
Рис. 126. Расчленение тела фаланги (Gale-odes): 1 — хелицера; 2 — педи-пальпа; 3 — ходильные конечности; 4— брюшко; 5— свободные задние сегменты груди; 6— пропельтидий; 7— глаза
а неблагоприятные последствия укусов связаны с попаданием в рану остающихся на хелицерах остатков пищи, в которых развиваются бактерии и образуется трупный яд. Хелицерами эти паукообразные пользуются также для рытья в почве норок, служащих их временными убежищами.
Педипальпы внешне походят на типичные ходильные конечности, но лишь в малой степени участвуют в беге и ходьбе; их вершины лишены коготков. Основная их функция — осязательная. Кроме того, сольпуги пользуются ими для захвата и удержания добычи, а также в общении с особями противоположного пола.
Последующие четыре пары ходильных конечностей головогруди состоят из семи члеников и несут на вершинах по паре коготков. Длинные и стройные, они фактически являются бегательными. Задние несколько мощнее передних, поскольку на них приходится нагрузка по удержанию массивного брюшка, а первая пара стройнее и короче других и выполняет главным образом осязательную функцию. При внимательном осмотре с брюшной стороны на тазиках и вертлугах последней пары ходильных конечностей могут быть обнаружены коло-
кольчикоподобные малеолы (привески). Предполагается, что они выполняют рецепторную функцию.
Подкласс Pseudoscorpiones — ложноскорпионы
Мелкие хелицеровые по внешнему виду напоминают скорпионов, но отличаются от последних тем, что брюшко их не имеет подразделения на передне- и заднебрюшие. Сходство этих паукообразных со скорпионами определяется клешневидными педипальпами. Характерно наличие паутинных желез. Дыхание трахейное. Развитие с метаморфозом. Хищники.
Для ознакомления с особенностями строения представителей подкласса лучше использовать не фиксированный материал, а постоянные тотальные препараты взрослых экземпляров книжного ложно-скорпиона.
Chelifer cancroides — книжный, или обыкновенный ложноскорпион
Материал. Лжескорпионы представляют собой группу широко распространенных, но ведущих скрытный образ жизни и поэтому малозаметных животных. Книжный ложноскорпион часто отмечается в библиотеках, книжных складах и иных книгохранилищах, а также чердаках, подвалах и других помещениях, где питается главным образом клещами пыли. В природных условиях эти животные встречаются в лесной подстилке, под отстающей корой деревьев, в дуплах и других подобных местообитаниях. Сбор достаточного количества экземпляров обычно затруднителен; наилучшие результаты может дать выгонка их из проб подстилки на фотоэклекторе. Анестезированных животных хранят в 70° этаноле, спиртовой материал вполне пригоден для изготовления постоянных препаратов.
Внешнее строение. Тело ложноскорпиона подразделяется на голо-вогрудной (просома) и брюшной (опистосома) отделы (рис. 127), визуально разграниченные бороздой. Брюшко шире груди и широко закруглено сзади, в результате чего тело имеет каплевидную форму. Оно в значительной мере уплощено, что позволяет этим членистоногим использовать щелевидные укрытия, такие как полости под отстающей корой или корешки книг. Головогрудь защищена сверху слитным головогрудным шитом. Брюшко состоит из 11 сегментов, границы между которыми различимы как на спинной, так и брюшной сторонах тела.
Хелицеры уложноскорпионов небольшие, клешневидные, сильно сближены между собой и направлены прямо вперед. На их вершинах
246
Тип Arthropoda
Класс Arachnida
247
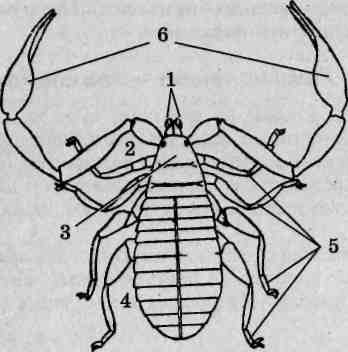
Рис. 127. Книжный, или обыкновенный, ложноскорпион (Chelifer cancroides): 1 — хелицеры; 2 — головогрудной отдел (просома); 3 — головогрудной щит; 4— брюшной отдел (опистосома); 5— ходильные конечности; 6— педипальпы
открываются протоки расположенных в головогруди парных паутинных желез. Педипальпы, вооруженные мощными клешнями, внешне напоминают таковые скорпионов, но снабжены длинными чувствительными волосками — трихоботриями. Педипальпами животные захватывают добычу, но ее обработка производится хелицерами. Остальные четыре пары головогрудных конечностей используются [ для ходьбы. На вершинах ходильных конечностей расположена пара коготков с небольшой присоской между ними. Эти приспособления j позволяют ложноскорпионам с легкостью перемещаться по вертикальным поверхностям (стволам деревьев, боковым поверхностям валунов, стенам и т.п.). Потревоженные животные пятятся либо двигаются боком, как это свойственно крабам и паукам-бокоходам. Расположенный на нижней стороне брюшка наружный половой аппарат имеет ; достаточно сложное строение, выяснение происхождения его частей представляет определенные затруднения.
Подкласс Aranei — пауки
Преимущественно мелкие и средних размеров хелицеровые, имеют слитные головогрудь и брюшко, соединенные стебельчатым пере-1
хватом. Хелицеры и педипальпы не клешнеобразные. Конечности последних двух сегментов брюшка преобразованы в паутинные бородавки. Жизнедеятельность пауков теснейшим образом связана с прядением паутины. Покровы тела образованы главным образом задуб-ленными белками, а не хитином. Дыхание легочное и трахейное. Развитие прямое. Все пауки — хищники.
На лабораторных занятиях могут быть использованы любые крупные представители таксона, но традиционным объектом является паук-крестовик.
Araneus diadematus — обыкновенный паук-крестовик
Материал. Этот паук в условиях лесной зоны Восточной Европы является одним из обычнейших представителей семейства пауков-круго-прядов (Araneidae). Для них характерно вертикальное расположение ловчих тенет, образованных концентрическими кругами паутины. В сумерки либо пасмурным днем крестовики располагаются по центру ловчей сети и их нетрудно собрать, подставляя снизу широкую пробирку либо иной сосуд. Хранят пауков в 80° этаноле.
Внешнее строение. Тело большинства пауков сильно укорочено и компактно. У крестовика между слитными головогрудью и брюшком находится стебелек (рис. 128, А), образованный первым брюшным сегментом. Головогрудь уплощена с боков, ее облегают нисходящие передний и боковые края головогрудного щита, на котором ближе к переднему краю размещены в два ряда четыре пары простых глаз, имеющих вид небольших черных точек.
Воспользовавшись ручной лупой или бинокуляром, можно ознакомиться с морфологией брюшной стороны головогруди (рис. 128, Б). По центру, между выпуклыми тазиками ходильных ног расположена пятиугольной формы умеренно склеротизированная пластинка — стер-нум. Спереди от стернума лежит полукруглой формы нижняя губа, сформировавшаяся из брюшной пластинки (стернита) II головогрудного сегмента. За ней следует пара челюстных лопастей, являющихся отростками тазиков конечностей этого же сегмента. Нижняя губа и челюстные лопасти окружают ротовое отверстие.
Хелицеры являются наиболее интенсивно склеротизированны-ми частями тела пауков. Они имеют фронтальное расположение; степень их жесткости можно установить нажатием препаровальной иглы. Хелицеры двучленистые; крупный основной членик имеет цилиндрическую форму и несет на вершине крючкообразный, подвижно прикрепленный дистальный членик. У крестовиков он выделяется своей темной окраской. Внутри него проходит открывающийся на вершине
248
Тип Arthropoda
Класс Arachnida
249
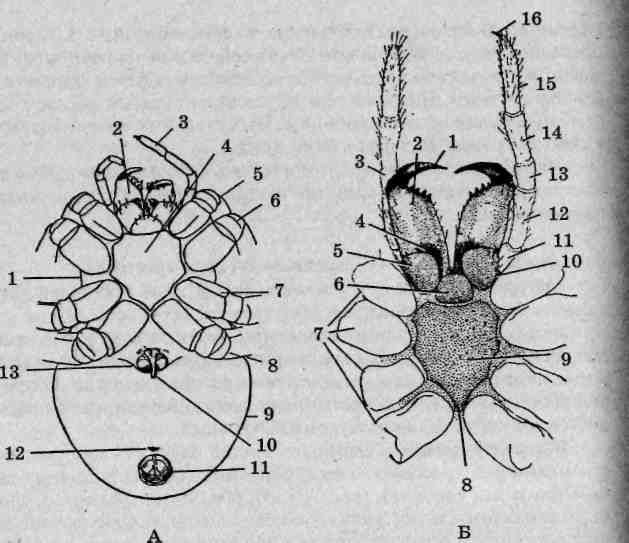
Рис. 128. Внешнее строение самки обыкновенного паука-крестовика (Am- \ neus diadematus): А — общий вид с брюшной стороны: 1 — головогрудь; 2-3 — головные конечности (2 — хелицера, 3 — педи-1 пальпа); 4 — стернум; 5-8 — грудные ходильные конечности (I-IV пары); 9—брюшко; 10— эпигене; 11 — паутинные боро- , давки; 12— стигмы трахей; 13— легочные крышечки; £—стро-: ение головогруди и расположение конечностей (хелицеры отогнуты вперед): 1 — когтевидный членик хелицеры; 2 — основной [ членик хелицеры; 3 — педипальпа; 4— пучок волосков челюстной лопасти; 5—челюстная лопасть; 6— нижняя губа; 7—тазики ног; 8 — пластинчатое расширение стернума; 9— стернум; ]0-i6— структурные части педипальпы (10— тазик; 11 — верт-Л луг; 12— бедро; 13— колено; 14— голень; 75— лапка; 16— ко- ; готок)
членика канал ядовитой железы. В состоянии покоя дистальный членик поджимается к поверхности основного членика. Благодаря заостренным вершинам и ядовитому секрету, хелицеры представляют собой серьезное оружие нападения и защиты. Пользуясь ими,
животные захватывают, удерживают и умерщвляют добычу, а также наносят раны, в которые затем впрыскивают содержащую пищеварительные ферменты слюну. Всем паукам свойственно внекишечное пищеварение.
Педипальпы выполняют главным образом осязательную функцию, ими животное ощупывает субстрат и окружающие предметы, фиксирует колебания нитей паутины. Как уже указывалось ранее, педипальпы являются единственными конечностями пауков, обладающими челюстными лопастями. У самок педипальпы по внешнему виду напоминают ходильные конечности. У самцов они укорочены и несут на вершинах специальные чашевидные расширения, с помощью которых во время спаривания собирается семенная жидкость и вводится в половые пути самки.
Ходильные конечности в числе четырех пар имеют типичное строение, несут на вершинах хитиновые коготки и используются для передвижения по различным субстратам, растительности или паутине. Характерной особенностью строения ходильных конечностей пауков является наличие дополнительного членика — колена, расположенного между бедром и голенью.
Округло-каплевидное брюшко прикреплено к головогруди стебельком, явственно смещенным вниз. Его дорсальная пластинка называется л ору мо м, авентральная — плагул ой. Дорсальная пластинка имеет большие размеры, нежели вентральная, и вследствие этого заворачивается на брюшную сторону. На спинной стороне брюшко крестовика имеет светлый крестообразный рисунок, послуживший основанием для названия животного. Сразу же за стебельком снизу размещается пара щелевидных стигмальных отверстий, ведущих в полость легочных мешков и прикрытых хитинизированными легочными крышечками. В местах расположения легочных мешков окраска покровов более светлая. Как и у скорпионов, легкие рассматриваются как впятившиеся вовнутрь дыхательные конечности. У самок между стигмами открывается женское половое отверстие, защищаемое характерной хитиновой пластинкой — эпигене. Ее форма у самок многих групп пауков видоспецифична и используется для их идентификации.
На конце брюшка, снизу имеется три пары небольших конических придатков. Этопаутинные бородавки, на вершинах которых открываются протоки паутинных желез. Бородавки первой и последней пар двучленистые, что указывает на их происхождение в результате преобразования брюшных конечностей. Бородавки средней пары,
250
Тип Arthropods
Класс Arachnids
251
 на
которые при осмотре из-за их мелких
размеров обращают внимание
в последнюю очередь, не обладают
членистостью и являются выростами
стенки тела. Между бородавками верхней
пары возвышается анальный
бугорок, на котором расположено анальное
отверстие.
на
которые при осмотре из-за их мелких
размеров обращают внимание
в последнюю очередь, не обладают
членистостью и являются выростами
стенки тела. Между бородавками верхней
пары возвышается анальный
бугорок, на котором расположено анальное
отверстие.
Подкласс Acari — клещи
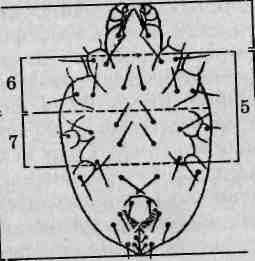
Мелкие, иногда микроскопических размеров хелицеровые. У взрослых особей деление тела на отделы обычно нечеткое. Для обозначения отдельных частей тела клещей применяется специальная терминология. В частности, в составе тела выделяют гнатосому, подосому, подразделяемую на проподосому и мезоподосому, а также опистосому. Гнатосома и проподосома в совокупности образуют протеросому, а метаподосома и опистосома — гистеросому (рис. 129). Дыхание трахейное или кожное. Развитие с метаморфозом, личинки имеют три пары ходильных конечностей, нимфы — четыре пары. В настоящее время клещей подразделяют на три отряда, с представителями двух из них целесообразно ознакомиться в ходе лабораторного практикума.
8
9
Рис. 129. Общая схема деления тела клещей на отделы: 1 — просома; 2— опистосома; 3— гнатосома; 4 — идиосома; 5— подосома; 6—проподосома; 7—метаподосома; 8— протеросома; 9—ги- стеросома \
Отряд Acariformes — акариформные клещи
Мелкие хелицеровые, тело взрослых особей подразделяется на протеросому (объединяет передние головогрудные сегменты) и гистеросому (объединяет два головогрудных и все брюшные сегменты),
граница между ними обычно неявна. Ротовой аппарат устроен просто, хелицеры грызущего или сосущего типа. Дыхание кожное или трахейное. Для индивидуального развития характерен анаморфоз. Эмбрионы с тремя парами зачатков конечностей. Сапрофаги, фитофаги, хищники или паразиты. Отряд подразделяется на два подотряда и более сотни семейств, объединяемых в серии.
Подотряд Sarcoptiformes — саркоптиформные клещи
Хелицеры грызущего типа. Почво- или растениеобитающие членистоногие, либо паразиты животных. Расселительную функцию выполняют взрослые особи.
Tyroglyphus farime — обыкновенный мучной клещ
Материал. Мучные клещи семейства Tyroglyphidae являются характерными вредителями запасов. При повышенной влажности хранящегося зерна, круп, муки они в массе могут развиваться в домах, зернохранилищах и складах. Другие тироглифоидные клещи обитают на сухофруктах, казеиновых отходах, сыре и многих других продуктах животного и растительного происхождения. На занятияхлучше воспользоваться постоянными тотальными препаратами. При использовании для их изготовления канадского бальзама клещей фиксируют 10% раствором формалина и окрашивают. При использовании гуммиарабико-вьгх заключающих сред Фора или Фора—Берлезе хранившихся в 70—80° этаноле клещей нужно обязательно провести через 40° спирт.
Внешнее строение. Взрослые мучные клещи имеют микроскопические размеры, длина тела самок не превышает 0,7 мм, самцов — 0,45 мм. Светлоокрашенное овальной формы тело Tyroglyphus farinae лишено видимых склеротизированных участков и одето тонкой хи-тиноидной кутикулой. Для рассмотрения особенностей внешней морфологии (рис. 130) лучше использовать более крупных самок, отличающихся менее сглаженными очертаниями тела.
Рассматривая клеща со спинной стороны, следует обратить внимание на бороздку, отделяющую треугольно-коническую протеросому от продолговатой гистеросомы. Спереди хорошо различимы направленные вперед клешневидные хелицеры. Глаз у этих паукообразных нет. На поверхности тела определенным образом распределены мелкие и крупные щетинки, в совокупности составляющие хетом.
Тип Arthropoda
Класс Arachnida
253

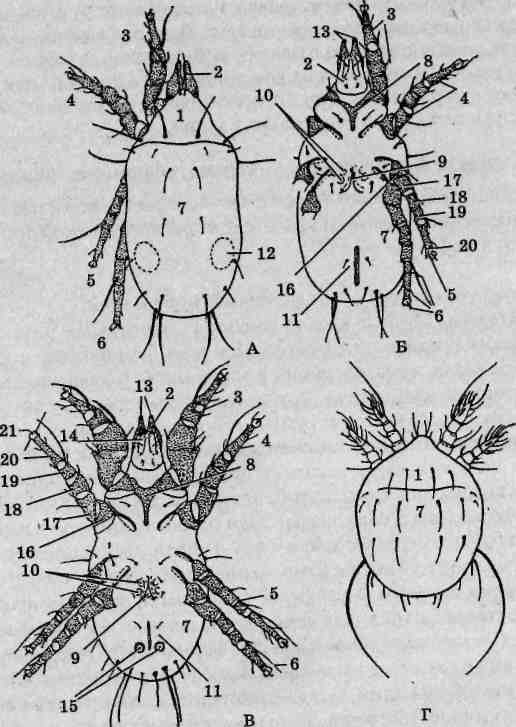
Рис 130. Обишо^ппъшщчпош^щ^Ту^рИиз/аппа^сш^ьт со спинной стороны (А), самка, вид сбрюшнои стороны (Б), са мец, вид с брюшной стороны (В), расселительная нимфа - га nonvc (ГУ 1 - протеросома; 2 - ротовые органы; 3-ь - i iv пар^ходильньххконе^остей; 7- гистеросома; *-ФУД™с; тинка (с^Гнум); 9- половое отверстие; 10- половые приемки 11 - анальное отверстие; 12- жировые железы, 13- мандибу лы; 74 - нижняя губа; 75 - анальные присоски; 76-27 -структурные части ходильной конечности (16- вертлуг• ч бед™ - колено; 19 - голень; 20 - лапка; 27 - коготок)
Схема размещения щетинок является важнейшим таксономическим признаком и используется при определении клещей отряда Acariformes. При изучении объекта с брюшной стороны (для этого выбирают другой, лежащий на спине, экземпляр либо аккуратно переворачивают препарат на предметном столике) на переднем конце тела видны направленные вперед продолговатые педипальпы. Между ними находится пластинчатая нижняя губа, несущая пару трехчленистых нижнегубных щупиков. Все четыре пары ходильных ног у самок примерно одинакового строения, у самцов первая пара конечностей отличается сильно утолщенными бедрами и коленными члениками. Все ходильные конечности оканчиваются изогнутым коготком. На уровне оснований задних ходильных конечностей (6-я пара) между двумя пластинчатыми клапанами располагается половое отверстие. Каждый клапан несет пару округлых половых присосок. В задней части брюшной стороны тела размещено анальное отверстие, имеющее виц длинной продольной щели. У самцов по бокам заднего конца тела расположена пара анальных присосок.
Подотряд Trombidiformes — тромбидиформные клещи
Хелицеры сосущего типа. Обитатели пресных водоемов, почво-и растениеобитающие членистоногие либо паразиты животных. Рассе-лительную функцию выполняют личинки.
Tetranychidae — паутинные клещи
Материал. Многие паутинные клещи родов Tetranychus, Schizo-tetranychus, Oligonychus, Panonychus, как и другие представители семейства Tetranychidae, являются массовыми вредителями культивируемых и иных растений. Особо благоприятны для их развития условия закрытого грунта с высокими температурами и низкой относительной влажностью воздуха. На интенсивное заселение растений паутинными клещами указывают локальное или точечное пожелтение листовых пластинок, других неодревесневших частей растений, которые при ближайшем рассмотрении оказываются затянутыми тонкой паутиной, что и послужило основанием для русскоязычного названия группы. В зимне-весенний период паутинные клещи могут быть собраны на оранжерейных или комнатных растениях. Во многих городах Белоруссии, европейской части России, Украины во второй половине лета липы мелколистная (Tilia cordald), широколистная (Tiliaplatyphyllos)
254
Тип Arthropoda
Класс Arachnida
255
 и
некоторые другие зачастую интенсивно
повреждаются липовым паутинным
клещом (Schizotetranychus
tiliarum). Подобно
молоди пауков, тетранихиды
на паутинках легко расселяются с
потоками воздуха. Поэтому
при демонстрации живых паутинных клещей
следует проявлять
осторожность, чтобы они случайно не
получили свободу и не колонизировали
имеющиеся в помещениях комнатные
растения. Для ознакомления
с внешней морфологией клещей лучше
использовать постоянные
препараты на основе заключающих сред
Фора или Фо-ра-Берлезе.
В случае использования материала,
хранившегося в
70-80° этаноле, его нужно обязательно
провести через 40° спирт.
и
некоторые другие зачастую интенсивно
повреждаются липовым паутинным
клещом (Schizotetranychus
tiliarum). Подобно
молоди пауков, тетранихиды
на паутинках легко расселяются с
потоками воздуха. Поэтому
при демонстрации живых паутинных клещей
следует проявлять
осторожность, чтобы они случайно не
получили свободу и не колонизировали
имеющиеся в помещениях комнатные
растения. Для ознакомления
с внешней морфологией клещей лучше
использовать постоянные
препараты на основе заключающих сред
Фора или Фо-ра-Берлезе.
В случае использования материала,
хранившегося в
70-80° этаноле, его нужно обязательно
провести через 40° спирт.
Внешнее строение. Взрослые паутинные клещи имеют микроскопические размеры, длина тела взрослых особей не превышает 0,5 мм. Форма тела овальная или продолговато-овальная (рис. 131). Самцы обычно несколько мельче и стройнее самок, с коротким телом, суживающимся к заднему концу. Спинная сторона тела выпуклая, брюшная — умеренно выпуклая. Прижизненная окраска чаще бледно-зеленая или | бледно-желто-зеленая. У взрослых особей обыкновенного паутинного клеща (Tetranychus urticae s.i), садового паутинного клеща (Schizotetranychus pruni), некоторых других видов по бокам тела видны характерной формы темные пигментные пятна. Летние самки красного плодового клеща (Panonychus ulmi) имеют ярко-красную окраску, тогда | какличинки, нимфы и самцы этого вида — зеленые. Впадающие в диа-паузу самки сливового паутинного клеща меняют бледно-желтую | окраску на лимонно-желтую.
Первичная сегментация тела утрачена, при взгляде сверху хорошо различима лишь граница между пропо до сомой иметаподосомой. На спинной стороне тела у представителей рода Tetranychus имеется участок со складчатой поверхностью покровов, образующих «ромбическую фигуру». Кожные покровы тела и конечностей несут большое количество щетинок, особенности хетома широко используются при определении таксономической принадлежности тетранихид. На спинной стороне по бокам проподосомы расположены простые глаза в виде карминно-красных точек. У большинства тетранихид их две пары.
Ротовой аппарат колюще-сосущего типа (рис. 132, А). Гнатосо-ма несетгипостом,стилофор состилетами ипарупедипальп (щупалец). Гипостом имеет вид четырехугольной пластинки, переходящей спереди в конический рострум,на вершине которого располагается ротовое отверстие. Образован гипостом слившимися базальными члениками педипальп. Стилофор является продуктом слияния пятых; члеников (голеней) хелицер, при взгляде сверху он имеет сердцевид-
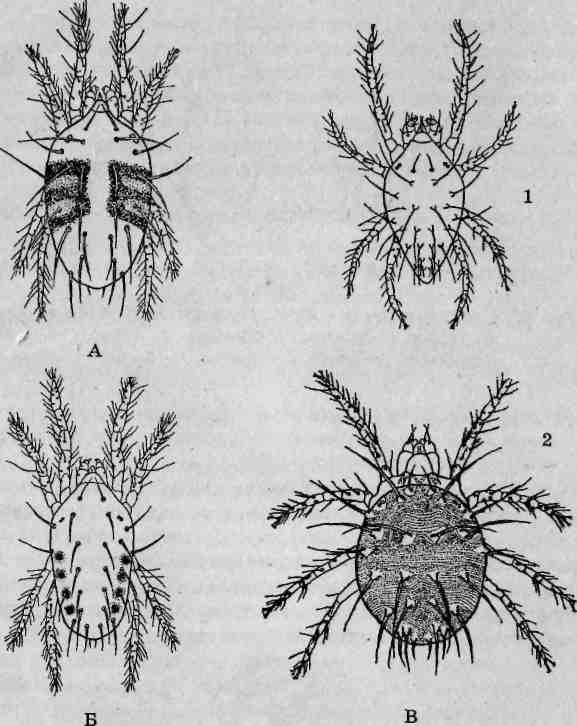
Рис. 131. Паутинные клещи {Tetranychidae): A — самка обыкновенного паутинного клеща (Tetranychus urticae si}; Б — самка садового паутинного клеща (Schizotetranychuspruni); В — самец (1) и самка (2) красного плодового клеща (Panonychus ulmi)
ную форму. Остальная дистальная часть хелицер видоизменилась в парные стилеты. С помощью такого ротового аппарата паутинные клещи способны высасывать содержимое растительных клеток и даже отдельных хлоропластов. Опустошенные клетки заполняются воздухом и при рассмотрении при увеличении могут обусловливать характерный
256 Тип Arthropoda
Класс Arachnids
257
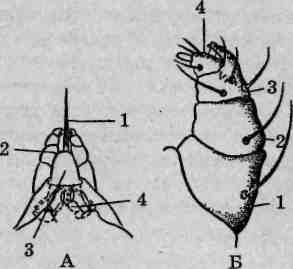
Рис. 132. Строение ротового аппарата паутинных клещей: А — гнатосома: 1 — стилет; 2 — пальпы; 3—стилофор; 4—козырек; Б — пальпа (щупальце): / — бедро; 2 — колено; 3 -голень; 4 — лапка
серебристый оттенок. Педипальпы (щупальца) паутинных клещей исходно шестичлениковые, но их тазики слились в гипостом, а остальная часть после слияния сегментов выглядит четырехчлениковой (рис. 132, Б).
Ходильные конечности паутинных клещей исходно шестичлениковые. В состав конечности входит тазик, неподвижно прикрепленный к корпусу тела, вертлуг, бедро, колено, голень и лапка (рис. 133), подвижно сочлененные друг с другом. Все они несут разнообразные щетинки, а лапка вооружена коготками. Как и большинство клещей, личинки тетранихид имеют три пары ходильных конечностей, с линькой на протонимфу появляется четвертая пара.
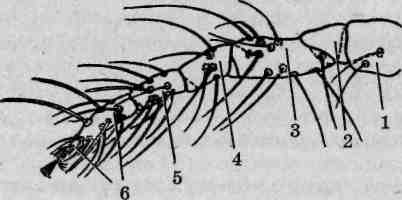
Рис. 133. Строение ходильной конечности паутинных клещей: 1 — тазик; 2— вертлуг; 3— бедро; 4— колено; 5— голень; 6— лапка
Летом самки откладывают яйца прямо в местах питания, осенью — в щелях коры, у основания почек. У ряда видов тетранихид (например, обыкновенного паутинного клеща) зимуют диапаузирующие самки. Из яиц отрождаются шестиногие личинки. Они превращаются в восьми-ногих протонимф, которые очень быстро линяют в дейтонимф. Взрослые особи, прежде чем приступить к размножению, должны пройти непродолжительный период дополнительного питания.
Отряд Parasitiformes — паразитоформные клещи
Мелкие и средних размеров хелицеровые, тело взрослых особей у большинства представителей лишено следов сегментации. Ротовой аппарат сложный, сосущего типа. Дыхание преимущественно трахейное. Эмбрионы с четырьмя парами зачатков конечностей. Хищники или паразиты животных, растительноядные формы отсутствуют. Переносчики возбудителей многих инфекционных заболеваний человека и животных.
В составе отряда выделяются три основные группы: гамазовые, иксодовые клещи и клещи-уроподы. На малом практикуме необходимо ознакомиться с отличающимися относительно крупными размерами и имеющими серьезное хозяйственное (эпидемиологическое, в частности) значение иксодовыми клещами.
Ixodes ricinus — собачий клещ
Материал. Клещи семейства Ixodidae характеризуются высокой степенью ландшафтно-биотопической специализации. Так, для таежной зоны Евразии характерен таежный клещ (Ixodes persulcatus), для зоны мелколиственных лесов — собачий клещ (Ixodes risinus), для степей — представители рода Dermacentor, полупустынь — Rhipicephalus, пустынь — Hyabmma. Собачий клещ наиболее многочислен в тех лесных массивах, где высока плотность копытных либо активно выпасается скот. Взрослых клещей собирают, окашивая сачком растительность на высоте 1,0-2,5 м либо применяя специальные экраны и иные приспособления. Личинок и нимф коллектируют, выполняя кошение по травостою и кустарникам. Иксодовые клещи являются эффективными переносчиками опасного заболевания человека — весенне-летнего клещевого энцефалита, имеющего вирусную этиологию, а также возбудителей ряда других болезней. Поэтому при сборе материала необходимо предпринять все необходимые меры предосторожности для
258
Тип Arthropods
Класс Arachnida
259
предотвращения укусов. Фиксировать клещей надо специальными составами: эфирно-спиртовой смесью (97 частей 20° этанола, 3 части ди-этилового (серного) эфира), жидкостью Удемса (87 частей 70° этанола, 8 частей ледяной уксусной кислоты, 5 частей глицерина) или Карнуа (6 частей 100° этанола, 3 части хлороформа, 1 часть ледяной уксусной кислоты). Это необходимо, чтобы предотвратить подгибание конечностей под подосому и их повышенную хрупкость при препаровке. На занятиях следует использовать тотальные микроскопические препараты взрослых самцов и самок, нимф и личинок. В качестве заключающей среды применяют канадский бальзам или жидкость Фора—Берлезе.
Внешнее строение. Иксодовые клещи имеют мешковидное тело, у непитавшихся особей сильно уплощенное в дорсовентральном направлении (рис. 134). У взрослых самок, нимф и личинок покровы тела весьма эластичны, имеются лишь сильно склеротизированные спинной и перитремальные щитки. Это позволяет им многократно увеличивать объем тела при питании. Самцы потребляют мало пищи и их тело защищено набором сильно хитинизированных щитков. Рассматривая препараты клещей под бинокуляром, следует обратить внимание на расположенные по бокам тела перитремальные щитки, на которых открываются дыхальца (стигмы) трахейной системы.
Ротовой аппарат колюще-сосущего типа (рис. 135). Устроен он сложнее, чем у тетранихид. Гнатосома, которую часто называют хоботком или головкой, несет специфически преобразованные хелицеры и педипальпы. Хелицеры имеют подвижные вершины (пальпы), вооружены режущими поверхностями и многочисленными загнутыми крючьями, способны выдвигаться вперед и втягиваться обратно. Производными педипальпявляются воротничок, гипос-томипальпы. Воротничок кольцом охватывает основание гнато-сомы. Снизу от него отходит вырост — гипостом, вооруженный большим числом шипиков. Сверху воротничок снабжен парой футляров (влагалищ), в обычном состоянии скрывающих основания хелицер. По бокам хелицеры несут четырехчлениковые щупики (пальпы). При питании клещ пробивает покровы тела животного-хозяина режущими поверхностями вершин хелицер и вводит ротовые части, находящиеся в сжатом состоянии. После их расхождения он надежно зая-коривается на хозяине. Необходимость преодоления зачастую весьма плотных защитных покровов хозяина заставляет клещей искать участки тела с более нежной кожей, например ушные раковины, веки,
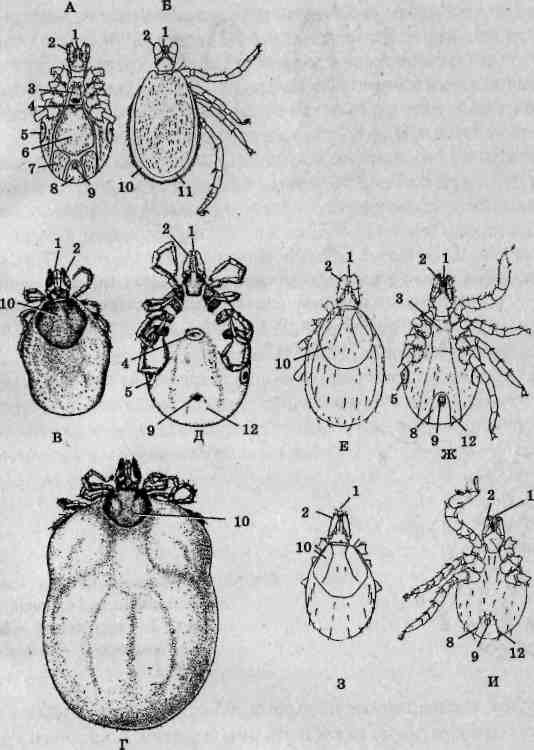
Рис. 134. Внешнее строение клеща Ixodes risinus: самца (А — вид сверху, Б — вид снизу); самки (В — непитавшаяся, вид сверху; Г— напитавшаяся, вид сверху; Д — слегка напитавшаяся, вид снизу); нимфа (Е — вид сверху, Ж — вид снизу); личинка (3 — вид сверху, И— вид снизу); / — гипостом; 2— педипальпы; 3— преге-нитальный щиток; 4— половое отверстие; 5 — перитремы с дыхальцами (стигмами); 6— срединный щиток; 7— аданальный щиток; 8— анальный щиток; 9— анальное отверстие; 10— спинной щиток; 11 — краевое поле; 12 — анальная бороздка
260
Тип Arthropoda
Класс Arachnida
261
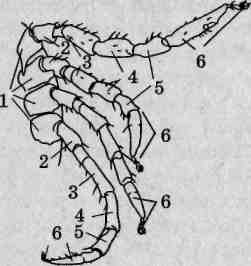
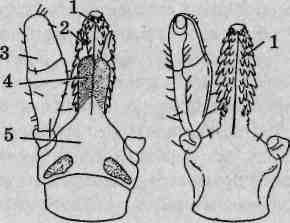
Рис. 135. Строение ротового аппарата иксодовых клещей: А — вид со спинной стороны; Б — вид с брюшной стороны; / — гипостом; 2 — дистальный членик хелицеры; 3 — пальпа; 4 — футляр для хелицер; 5 — воротничок
Рис. 136. Строение ходильных конечностей иксодовых клещей: / — тазик; 2 — вертлуг; 3 — бедро; 4 — колено; 5 — голень; 6 — лапка
Конечности иксодовых и тетраниховых клещей имеют схожее строение (рис. 136). Личинки обладают тремя парами ходильных конечностей, нимфы — четырьмя. Развитие иксодид протекает аналогичным образом, однако имеется лишь одна нимфальная стадия. Не имея глаз, клещи взбираются на возвышенные или выступающие предметы (вершины былинок, веточек кустарников, выступающих над открытыми участками ветвей деревьев) и неподвижно располагаются с вытянутыми вперед передними конечностями в ожидании жертв. Соприкосновение с предметами, фактура поверхности которых напоминает шерсть, вызывает соответствующую реакцию фиксации этих животных. Прокормителями личинок Ixodes ricinus обычно становятся мышевидные грызуны, нимф -* зайцы и ежи, тогда как взрослых особей — копытные, хищные псовые (собаки, волки, лисы), а также человек. Соответствует этому и распределение клещей по ярусам растительности: личинки, как правило, не поднимаются выше травяного яруса, нимфы размещаются на низкорослых кустарниках, взрослые стерегут жертв на высоких кустарниках и ветвях нижнего яруса крон. Таким образом, они выбирают те места, где максимальна возможность встречи с адекватными видами жертв.
подбородок, подмышечные и паховые области. Собачьи клещи в норме питаются однократно на каждой стадии развития, после чего приступают к подготовке линьки или откладке яиц. Питание иксодовых клещей является сложным и длительным (многочасовым и более) процессом и не сводится к простому поглощению пищи: в течение этого периода интенсивно протекают рост и развитие организма. Попытки механическим усилием удалить эктопаразита с места прикрепления часто оканчивается разрывом тела. При этом гнатосома остается в тканях хозяина, что может повлечь для него самые неприятные последствия (от образования нарыва до заражения крови).
Класс Asteroidea
263
Тип Echinodermata — иглокожие
Иглокожие — единственные представители вторичноротых, рассматриваемые на занятиях данного практикума. Радиальная симметрия тела у них вторична, в той или иной степени развиты внутренний известковый скелет и амбулакральная (воднососудистая) система. На эти особенности строения иглокожих следует обратить особое внимание при выполнении лабораторных работ.
Иглокожие являются обитателями морей и весьма требовательны к уровню солености воды. Содержание их в аквариумах с морской водой весьма затруднительно. Все это осложняет знакомство с представителями типа и обеспечение занятий необходимым материалом. Целесообразно поэтому ограничиться непосредственным ознакомлением с отдельными особенностями строения представителей двух классов иглокожих: морских звезд {Asteroidea) и морских ежей (Echinoidea).
Класс Asteroidea — морские звезды
Материал. Классическим объектом для знакомства с представителями таксона является морская звезда Asterias rubens — обычный обитатель морских мелководий Северной Атлантики, Баренцева и Белого морей. Ближе к берегу эти животные подходят в середине лета. Размах лучей крупных экземпляров может достигать 30 см; окраска, вопреки названию, разнообразна, но ограничивается красно-коричневой гаммой. В дальневосточных морях достаточно обычны имеющие чаще всего фиолетовую окраску Asterias amurensis, а также образующие крупные скопления патирии (Patiria). Последних можно использовать для ознакомления с особенностями внешнего строения морских звезд, но они малопригодны для анатомирования в учебных целях.
Морских звезд следует хранить в 70° этаноле. Хранение в формалине ведет к разрушению скелетных элементов, плохо в нем сохраняется и большинство внутренних органов. Собранных животных погружают целиком в этанол указанной крепости. Затем на крупных экземплярах следует сделать надрезы и выпустить полостную жидкость, заменив ее этанолом. Важно проследить, чтобы в итоге крепость спирта осталась на уровне, обеспечивающем сохранность объектов.
Внешнее строение. Тело Asterias rubens уплощено в дорсовен-тральном направлении и очертаниями повторяет пятиконечную звезду (рис. 137, А). Выделяются центральный диск и пять отходящих от него лучей, называемых также руками. Более широкие у основания,
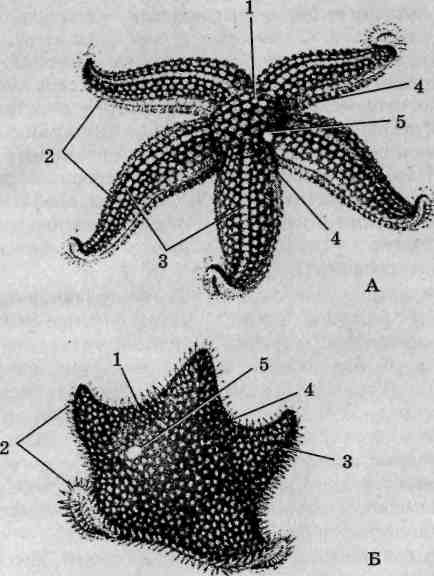
Рис. 137. Внешнее строение морских звезд: Asterias (А) и Patiria (Б); вид с аборальной стороны: 1 — центральный диск; 2 — лучи (руки); 3 — радиусы; 4 — интеррадиусы; 5— мадрепоровая пластинка
лучи сужаются к вершинам. У крупных экземпляров они зачастую бывают различной длины, что свидетельствует об имевших место травмах, компенсированных благодаря высокой регенерационной способности этих животных. Для облегчения описания морфологии иглокожих используют понятия интеррадиусов и радиусов. Радиусы — это линии, соответствующие лучам симметрии морской звезды, интеррадиусы соединяют центр диска с точками соединения оснований смежных лучей.
Ротовое отверстие расположено в центре диска с нижней брюшной стороны тела. Последняя уплощена и носит название оральной (ротовой), или амбулакральной. На ней вдоль осевой линии каждого луча от основания до вершины проходит амбулакральная бороздка,
264
Тип Echinodermata
Класс Asteroidea
265
несущая четыре ряда амбулакральных ножек — цилиндрических, со-сочковидных выростов. С помощью ручной лупы нетрудно рассмотреть, что их вершины снабжены небольшими дисковидными присосками. Эти ножки отличаются от остальных участков амбулакральной поверхности окраской, тонкими покровами и легко прогибаются при нажатии препаровальной иглой. Именно с их помощью морские звезды передвигаются по дну и осуществляют различные манипуляции лучами. На вершине каждой амбулакральной бороздки расположена непарная чувствительная амбулакральная ножка, снабженная химическими рецепторами и несущая у основания светочувствительное пигментное пятно — глазок. Обнаружить его у фиксированных экземпляров достаточно трудно.
Верхняя, аборальная, или антиамбулакральная сторона тела гораздо более выпуклая, нежели оральная. В центре диска расположено с трудом различимое поровидное анальное отверстие. Водном из интеррадиусов имеется небольшая ситовидная, или мадрепо-ровая пластинка, которая названа так из-за сходства в строении с фрагментами скелета мадрепоровых кораллов. Она выполняет функцию своеобразного керамического фильтра, через который морская вода поступает в амбулакральную систему животного. Мадрепо-ровую пластинку следует рассмотреть с помощью ручной лупы либо бинокуляра, а легкое царапание препаровальной иглой дает представление о механических свойствах этого образования.
Тело морской звезды одето мягкой тонкой кожей. На поверхности покровов нетрудно обнаружить образования трех типов: выступающие из кожи тупые известковые иглы и пинцетовидные либо клювовидные педицеллярии, а также конические тонкостенные выросты — папулы, выполняющие функции кожных жабр. Педицеллярии имеются только у педицелляриевых звезд, к которым и принадлежит Asterias rubens. Животное использует их главным образом для очистки поверхности тела от загрязнения оседающими на нее частицами.
Под кожей морской звезды залегает сплошным слоем наборный пластинчатый известковый скелет. Такое его строение обеспечивает свободу движений лучей животного. Скелет носит название наружного кожного, но имеет мезодермальное происхождение. В лучах Asterias rubens с оральной стороны расположение скелетных пластинок строго упорядочено (рис. 138), чего нельзя сказать о центральном диске либо обо всем теле многих других представителей класса, например патирий. Вдоль срединной оси каждой руки, образуя дно амбулакральной бороздки, проходят два ряда амбулакральных скелетных пластинок.
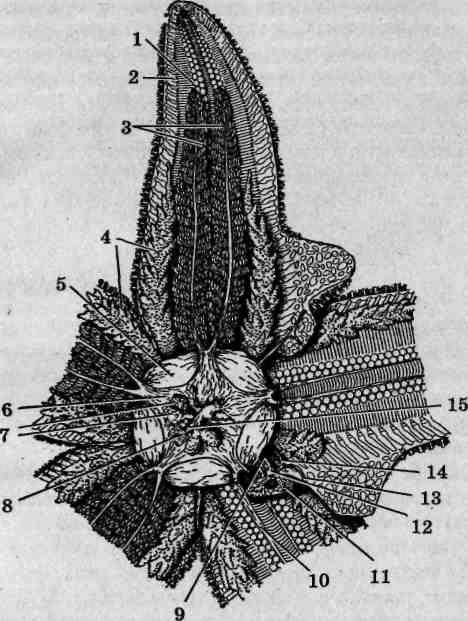
Рис. 138. Вскрытая морская звезда Asterias rubens: 1 — амбулакральные пластинки; 2 — маргинальные пластинки; 3 — печеночные придатки (мешки); 4— гонады; 5— оральный отдел желудка; 6— аборальный отдел желудка; 7— ректальные железы; 8— фрагмент спинной стенки тела с анальным отверстием; 9— каменистый канал; 10 — мускулы-ретракторы желудка; 11 — участок кожи с мадрепоровой пластинкой; 12 — стенка осевого синуса; 13— половой столон; 14— половой проток; 15— задняя кишка
От прочих они отличаются еще и тем, что насквозь пронизаны тонкими поровыми канальцами, соединяющими амбулакральные ножки с их внутренними ампулами. По сторонам к рядам амбулакральных примыкает по ряду адамбулакральных пластинок. Далее
266
Тип Echinodermata
Класс Asteroidea
267
следует по ряду интеррадиальных пластинок и по два ряда нижних маргинальных (краевых) пластинок. В интеррадиусах по краям ротового отверстия расположены так называемые зубы, представляющие собой заостренные отростки краевых адамбулакральных пластинок. Несмотря на подобное название, их роль в обработке пищи незначительна. Скелет аборальной стороны луча образован либо немногими рядами скелетных пластинок, либо многочисленными тонкими известковыми перекладинами. Знакомиться с расположением скелетных пластинок лучше на сухих крупных экземплярах Asterias rubens, у которых предварительно с одного из лучей слущены амбулакральные ножки и кожные покровы. При отсутствии такого материала скелетные элементы целесообразно рассмотреть после вскрытия фиксированных морских звезд.
Вскрытие морской звезды. При наличии в достаточном количестве раздаточного материала весьма желательно, чтобы вскрытие было выполнено каждым обучающимся. В противном случае проводят демонстрационное вскрытие либо для знакомства с особенностями внутреннего строения используют хранящиеся в фиксаторе уже вскрытые крупные экземпляры.
Отыскав на аборальной стороне тела мадрепоровую пластинку, начинают вскрытие животного с помощью анатомических ножниц. Разрез ведут по боковой стороне тела, несколько смещая его к аборальной. Мадрепоровую пластинку огибают по аборальной стороне, не достигая центра диска. Центральный участок диска, несущий анальное отверстие, также лучше обогнуть краевым разрезом. Все операции легче выполнять, держа животное в руках. Закончив разрез, звезду помещают в препаровальную ванночку оральной стороной вниз и закрепляют булавками. Затем осторожно снимают кожные покровы аборальной стороны тела, отгибая их и подсекая удерживающие связки, в результате чего открывается полость тела морской звезды.
Внутреннее строение. Описанная техника вскрытия позволяет вначале ознакомиться с особенностями строения пищеварительной системы. В пределах центрального диска дорсально по интеррадиусам залегают гроздьевидные ректальные железы, являющиеся выростами задней кишки. Как и часть стенок мешковидного желудка, они зачастую удаляются вместе с кожными покровами аборальной стороны тела. Занимающий центральное положение (рис. 138) объемистый складчатый желудок может содержать непереваренные остатки пищи. Однако Asterias rubens, как и другие представители семейства
Asteridae, весьма широко использует возможность выворачивать через ротовое отверстие часть желудка и переваривать пищу, фактически не заглатывая ее. От желудка в каждый луч отходит непарный проток, переходящий в протоки парных бурых печеночных придатков (мешков), которые почти целиком заполняют внутреннее пространство луча неполовозрелых особей. Основная их функция — продуцирование пищеварительных ферментов. При постепенном удалении печеночных желез и желудка можно проследить, как последний сужается к ротовому отверстию, переходя в короткий пищевод. При этом следует подсечь мускулы-ретракторы, обеспечивающие втягивание вывернутой через ротовое отверстие части желудка.
Половая система. Парные перистые гонады расположены в интеррадиусах, продолжаясь в соответствующие лучи. Их размеры зависят от степени полового созревания и могут сильно варьировать — от малозаметных структур в пределах центрального диска до хорошо развитых образований, заполняющих большую часть полости лучей. В интеррадиусах на аборальной стороне тела они открываются половыми отверстиями, практически неразличимыми на фиксированном материале. Морские звезды раздельнополые животные, но женские и мужские половые железы внешне неразличимы. Половая принадлежность особей может быть установлена только по результатам исследования продуцируемых половых продуктов.
Амбулакральная (воднососудистая) система начинается мадрепо-ровой пластинкой, канальцы которой ведут в S-образно извитый каменистый канал, входящий в осевой комплекс органов. Последний спускается с аборальной стороны центрального диска к оральной. Другие компоненты осевого комплекса неразличимы при используемой технике фиксации и анатомирования объекта. Стенки каменистого канала обызвествлены, что легко выявить простым нажатием пинцета. Каменистый канал впадает в кольцевой канал амбулакральной системы, опоясывающий пищевод. Сам канал обычно неразличим, но его месторасположение может быть установлено по темно-желтым или светло-бурым размером со спичечную головку тидемановым тельцам (железам). Они располагаются в интеррадиусах попарно и выполняют функции лимфоцитарных органов. Общее число тидемановых телец равно девяти, поскольку на месте расположения одного из них в кольцевой канал амбулакральной системы впадает каменистый канал. Вырабатываемые тидемановыми тельцами амебоидные клетки обеспечивают удаление поглощаемых ими продуктов метаболизма и иных ненужных и опасных для организма соединений, т.е. выполняют выделительную и защитную функции.
268
Тип Echinodermata
Класс Echinoidea
269
Если аккуратно удалить из луча печеночные придатки (при длительном хранении материала они могут мацерироваться и разрушаться при прикосновении, превращаясь в кашеобразную массу), то под ними обнажается внутренняя сторона амбулакральной бороздки. По ней проходит радиальный канал амбулакральной системы, дающий ответвления к ее ампулам, располагающимся здесь в четыре ряда. Они представляют собой продолговатые мешочки с эластичными, мускулистыми стенками. Воднососудистая система заполнена жидкостью, основой которой является поступающая через мадрепоровую пластинку морская вода. Сокращение ампул приводит к растяжению ножек. Прикрепившись присосками, последние сокращаются, нагнетая жидкость в ампулы. Одновременно животное подтягивается к месту прикрепления присосок амбулакральных ножек. В связи с отсутствием в лучах выраженных пучков мышечных волокон основными функциями амбулакральной системы являются изменение формы тела и обеспечение передвижения животных. В некоторой степени водно-сосудистая система участвует в осуществлении дыхания и выделения.
Нервная система у морских звезд представлена тремя радиаль-но-симметричными системами, залегающими в теле животного на разных уровнях. Проще всего ознакомиться с залегающей поверхностно, непосредственно под кожными покровами аборальной стороны тела аборальной (апикальной) нервной эндоневральной системой. Она выявляется при внимательном рассмотрении внутренней поверхности ранее удаленной аборальной стенки тела в виде пяти бурых радиальных тяжей, от центра диска уходящих в лучи. Эктонев-ральная (оральная, адоральная нервная) система обнажается после полного выщипывания амбулакральных ножек и представляет собой пять бурых тяжей, идущих в лучи по дну амбулакральных бороздок от околоротового нервного кольца. Залегающая в толще тела гипоневральная система при описанной технике фиксации и препарирования объекта выделена быть не может.
Кровеносная и перигемальная системы остаются за рамками рассмотрения, поскольку требуют иной техники препаровки и анатомирования.
Специальных органов выделения у иглокожих нет. Эту функцию выполняют амебоидные клетки полостной жидкости.
Органами дыхания служат кожные жабры — короткие тонкостенные выпячивания стенки тела (папулы), способные к сокращению. Они могут окружать кольцами скелетные иглы и хорошо видны при рассмотрении под бинокуляром верхней стенки тела морской звезды.
Класс Echinoidea — морские ежи
Материал. Наиболее доступным объектом для знакомства с представителями данного класса являются морские ежи рода Strongylocentrotus, например массовый обитатель литорали Баренцева, Белого и других северных морей Strongylocentrotus droebachiensis. Животные обитают на глубине свыше 5 м, что затрудняет их добычу. Техника фиксации морских ежей аналогична описанной выше для морских звезд.
На лабораторных занятиях целесообразно ограничиться 'знакомством с внешним видом, строением скелетного панциря и его игловидных придатков, ротового аппарата и педицеллярий морского ежа. Для этого используются фиксированный или высушенный материал, смонтированные на картонках или в застекленных коробках распиленные панцири ежей, а также постоянные микроскопические препараты их педицеллярий.
Внешнее строение. Морской еж имеет форму сплюснутого шара (рис. 139), тело плотно усеяно подвижно сочлененными крупными обызвествленными иглами. Наиболее крупные из них носят название первичных, более мелкие — вторичных, тогда как самые мелкие — миллиарных.
Ротовой полюс располагается на более уплощенной стороне тела. Как и морские звезды, морские ежи передвигаются по горизонтальным поверхностям ротовым полюсом вниз. При рассмотрении фиксированных экземпляров можно заметить, что расположенный здесь рот окружает кожистое поле, называемое перистомальным (ротовым), посередине которого располагаются торчащие наружу острые
Рис. 139. Морской еж Echinocidaris с сегментом поверхности, освобожденной от игл: 1 — интерамбулакральные ряды пластинок; 2 — амбулакральные ряды пластинок
270
Тип Echinodermata
Класс Echinoidea
271
долотообразные зубы. С противоположной стороны в окружении коротких мелких игл размещается анальное поле (перипрокт). Несколько в стороне от его центра открывается анальное отверстие, выявление которого затруднительно. Среди игл расположены короткие подвижные придатки, несущие на вершинах пинцетоподобные или клювовидные образования — характерные для иглокожих педицел-лярии.
Строение скелетного панциря. Скелет морских ежей имеет форму уплощенной с полюсов сферы (рис. 140), образованной неподвижно спаянными между собой известковыми пластинками. Выделяются два основных их типа. Более мелкие амбулакральные пластинки пронизаны тонкими сквозными отверстиями (эта особенность строения выявляется при внимательном рассмотрении высушенных панци-
Рис. 140. Схема строения скелета морских ежей, вид с оральной стороны: 1 — амбулакральные ряды пластинок; 2— интерамбулакральные ряды пластинок; 3— ротовое отверстие; 4— зубы; 5— оральная мембрана (перистом); 6— оральные пластинки с порами
рей); более крупные интерамбулакральные пластинки цельные. Пластинки каждого типа образуют меридионально располагающиеся чередующиеся парные ряды, общее их число равно десяти.
Скелетные пластинки имеют форму пятиугольников. Граница между соседними меридиональными рядами амбулакральных и интерамбулакральных (т.е. разноименных) пластинок имеет вид прямой линии, тогда как между одноименных — зигзагообразной. Принято считать, что ряды амбулакральных пластинок соответствуют рад иусам, а интерамбулакральных — интеррадиусам. Таким образом, здесь выделяются все те же пять радиусов и пять интеррадиусов.
Каждая пара рядов амбулакральных пластинок заканчивается у анального полюса генитальной пластинкой, несущей наружное половое отверстие. Всего таких пластинок четыре, так как пятая, имеющая несколько более крупные размеры и пронизанная множеством мелких отверстий, представляет собой мадрепоровую пластинку. Генитальные пластинки чередуются с более мелкими глазными (окулярными), каждая из которых пронизана отверстием специализированной амбулакральной ножки, несущей глазок. Прочие амбулакральные пластинки несут четыре-пять пар отверстий; их считают сложными, то есть образованными в результате слияния соответствующего количества простых пластинок.
Наружная поверхность панциря усеяна множеством бугорков, к которым шарнирно причленяются известковые иглы.
Строение ротового аппарата. На оральной стороне тела наружу выступают пять радиально-симметрично расположенных острых долото-видных зубов. Они являются частью сложно устроенного жевательного аппарата, который называется аристотелевым фонарем. В его состав входит 25 известковых скелетных элементов (перекладин и пластинок), подвижно сочлененных как между собой, так и с остальным панцирем и снабженных мускулатурой. Долотовидные зубы являются видимой снаружи частью аристотелева фонаря, а просвет между зубами — ротовым отверстием. Ежи рода Strongylocentrotus питаются живыми обрастаниями каменистых поверхностей (водорослями и пр.), соскребая и измельчая их с помощью жевательного аппарата.
Строение педицеллярий морского ежа. Педицеллярии разбросаны между иглами по всему телу морского ежа. По своему происхождению они являются модифицированными иглами. Педицеллярия состоит (рис. 141) из подвижного стебелька, снабженного известковой опорной осью, гибкой шейки и головки, образованной тремя смыкающимися
272
Тип Echinodermata


Рис. 141. Педицеллярии морского ежа Strongylocentmtus droebachiensis: А — шароносная; Б — змееголовая; В — трилистная; Г— трезубая; 1 — стебелек (стержень); 2 — шейка; 3 — головка; 4 — створки
створками, работающими наподобие щипцов или пинцета. Со строением этих образований можно ознакомиться, воспользовавшись соответствующими постоянными микроскопическими препаратами.
Основное назначение педицеллярии — очистка поверхности тела, промежутков между иглами от экскрементов, оседающих частиц ила и других загрязнений. Педицеллярии могут захватывать прикоснувшихся к ним мелких животных, умерщвлять их и передавать, как в эстафете, другим педицелляриям и далее к ротовому отверстию. Кроме того они, могут травмировать мягкие ткани врагов морских ежей и отрывать педицеллярии хищных морских звезд. У ряда представителей класса есть специализированные ядовитые педицеллярии, снабженные ядовитыми железами, которые выполняют главным образом защитные функции.