

Гликгенолиз.
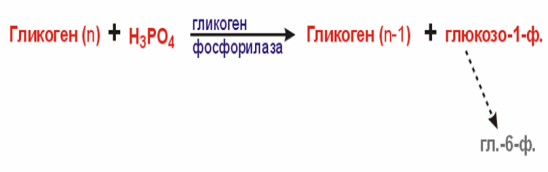
В результате фосфоролирования происходит разрыв гликозидных связей. Гликогенфосфорилаза - димерная молекула.
Вопрос№25 Константа Михаэлиса.
Михаэлиса
константа,
один из важнейших параметров кинетики
ферментативных реакций, введенный
немецкими учеными Л. Михаэлисом (L.
Michaelis) и М. Ментен в 1913; характеризует
зависимость скорости ферментативного
процесса от концентрации субстрата.
Согласно теории Михаэлиса — Ментен,
первым этапом любого ферментативного
процесса является обратимая реакция
между ферментом (Е)
и субстратом (), приводящая к образованию
промежуточного фермент-субстратного
комплекса (ES),
который затем подвергается практически
необратимому расщеплению на продукт
реакции (Р)
и исходный фермент:
 Реакции
образования и распада комплекса ES
характеризуются
константами скорости k1,
k-1,
k2.
Если концентрация субстрата значительно
превышает концентрацию фермента (S)
>> (E))
и, следовательно, концентрация ES
становится постоянной, скорость
ферментативной реакции (u) выражается
уравнением:
Реакции
образования и распада комплекса ES
характеризуются
константами скорости k1,
k-1,
k2.
Если концентрация субстрата значительно
превышает концентрацию фермента (S)
>> (E))
и, следовательно, концентрация ES
становится постоянной, скорость
ферментативной реакции (u) выражается
уравнением:
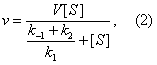 где
— максимальная скорость реакции,
достигаемая при полном насыщении
фермента субстратом. Соотношение
констант скорости
где
— максимальная скорость реакции,
достигаемая при полном насыщении
фермента субстратом. Соотношение
констант скорости
![]() также
является константой (Кm),
получившей название Михаэлиса
константа
Подставляя в уравнение (2) Михаэлиса
константа,
получаем уравнение Михаэлиса —
Ментен:
также
является константой (Кm),
получившей название Михаэлиса
константа
Подставляя в уравнение (2) Михаэлиса
константа,
получаем уравнение Михаэлиса —
Ментен:
![]() Из
уравнения (3) следует, что Михаэлиса
константа
численно равна концентрации субстрата,
при которой скорость реакции составляет
половину максимально возможной (см.
рис.).
В ряде случаев, когда величина k1
мала и ею можно пренебречь, Михаэлиса
константа
становится равной
Из
уравнения (3) следует, что Михаэлиса
константа
численно равна концентрации субстрата,
при которой скорость реакции составляет
половину максимально возможной (см.
рис.).
В ряде случаев, когда величина k1
мала и ею можно пренебречь, Михаэлиса
константа
становится равной
![]() и
может служить мерой сродства субстрата
к ферменту. Михаэлиса
константа
имеет размерность концентрации.
Практически величину Михаэлиса
константа
находят различными графическими
методами, исследуя зависимость скорости
ферментативной реакции от концентрации
субстрата.
и
может служить мерой сродства субстрата
к ферменту. Михаэлиса
константа
имеет размерность концентрации.
Практически величину Михаэлиса
константа
находят различными графическими
методами, исследуя зависимость скорости
ферментативной реакции от концентрации
субстрата.
Вопрос№26Обратимые и необратимые ингибиторы ферментов. Характерные черты конкурентного, неконкурентного и бесконкурентного ингибирования.
Ферментативный ингибитор — вещество, замедляющее протекание ферментативной реакции. Различают обратимые и необратимые ингибиторы. Обратимые делятся на конкурентные, неконкурентные, бесконкурентные.
Обратимое ингибирование
В
 этом случае ингибитор связывается в
активном
центре
фермента и конкурирует за него с
субстратом.
Таким образом, конкурентный ингибитор
не связывается с фермент-субстратным
комплексом (ES на рис.1), то есть константа
диссоциации Ki'
>> 1.
этом случае ингибитор связывается в
активном
центре
фермента и конкурирует за него с
субстратом.
Таким образом, конкурентный ингибитор
не связывается с фермент-субстратным
комплексом (ES на рис.1), то есть константа
диссоциации Ki'
>> 1.
Конкурентный ингибитор обычно структурно схож с субстратом, однако фермент не способен катализировать реакцию в присутствии ингибитора из-за отсутствия у последнего необходимых функциональных групп.Схема конкурентного
ингибирования и уравнение Михаэлиса-Ментен для него выглядят следующим образом:
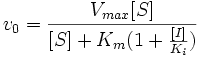
Видно, что при конкурентном ингибировании максимальная скорость реакции Vmax не меняется, а кажущаяся константа Михаэлиса увеличивается в (1 + [I]/Ki) раз. Поэтому в двойных обратных координатах Лайнуивера-Берка (зависимость 1/v0 от 1/[S]) при разных концентрациях ингибитора получают семейство прямых с различным наклоном, пересекающихся в одной точке на оси ординат.Константу ингибирования Ki обычно определяют так: проводят ряд измерений кажущейся константы Михаэлиса при различных концентрациях ингибитора, затем строят зависимость этой величины от концентрации ингибитора. Тангенс угла наклона полученной прямой равен Km/Ki.
Неконкурентное ингибирование
Неконкурентный ингибитор не мешает связыванию субстрата с ферментом. Он способен присоединяться как к свободному ферменту, так и к фермент-субстратному комплексу с одинаковой эффективностью. Ингибитор вызывает такие конформационные изменения, которые не позволяют ферменту превращать субстрат в продукт, но не влияют на сродство фермента к субстрату.
Схема и уравнение Михаэлиса-Ментен в случае неконкурентного ингибирования:
|
|
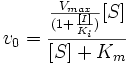
При неконкурентном ингибировании константа Михаэлиса не изменяется, а максимальная скорость реакции уменьшается в (1 + [I]/Ki) раз. Поэтому в двойных обратных координатах семейство прямых, отвечающих разным концентрациям ингибитора, пересекается в одной точке на оси абсцисс.
Бесконкурентное ингибирование
П
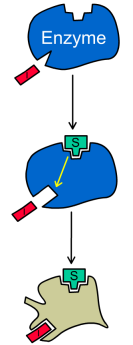 ри
бесконкурентном ингибировании ингибитор
связывается только с фермент-субстратным
комплексом, но не со свободным ферментом.
Субстрат, связываясь с ферментом,
изменяет его конформацию, что делает
возможным связывание с ингибитором.
Ингибитор, в свою очередь, так меняет
конформацию фермента, что катализ
становится невозможным.
ри
бесконкурентном ингибировании ингибитор
связывается только с фермент-субстратным
комплексом, но не со свободным ферментом.
Субстрат, связываясь с ферментом,
изменяет его конформацию, что делает
возможным связывание с ингибитором.
Ингибитор, в свою очередь, так меняет
конформацию фермента, что катализ
становится невозможным.
Схема и уравнение Михаэлиса-Ментен в случае бесконкурентного ингибирования:
|
|
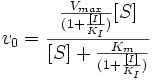
Максимальная скорость реакции и кажущаяся константа Михаэлиса уменьшаются в одинаковое число раз. Поэтому в двойных обратных координатах для разных концентраций субстрата получаем семейство параллельных прямых.