

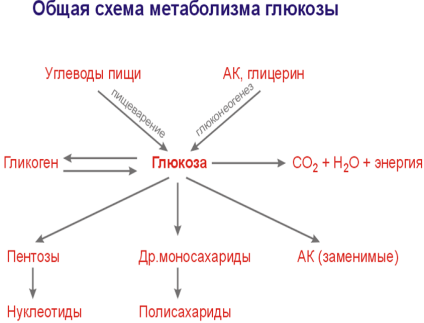
69. Глюкоза как важнейший метаболит углеводного обмена. Источники и пути использования глюкозы в организме.

Транспорт глюкозы из крови в клетку зависит от инсулина. Кроме инсулинонезависимых тканей – мозг и печень. Скорость поступления зависит от концентрации глюкозы в плазме крови.
ГЛЮТ – глюкозные транспортеры.
Первичное химическое превращение глюкозы в клетках - фосфорилирование – результат взаимодействия с АТФ → глюкоза-6-фосфат (активированная форма моносахаридов). Г-6-Ф не может свободно выйти из клетки. В результате фосфорилирования глюкоза «запирается» в клетке. В паренхиматозных клетках печени есть 2 фермента, катализирующих эту р-ю (из глюк. – глю.-6-фосф.) – гексокиназа и глюкокиназа (в др. органах –только гексокиназа).
1 фермент – гексокиназа Km<0,1 ммоль/л (сродство к глюкозе очень высоко), следовательно, max скорости реакции достигается при низк. конц. глюк. Глюкоза-6-фосфат ингибирует гексокиназу.
2 фермент - глюкокиназа – фермент в печени. Высокое значения Km ~ 10 ммоль/л.
Вопрос 73. Соотношение превращений субстратов и процессов, происходящих в печени, мышцах и жировой ткани.
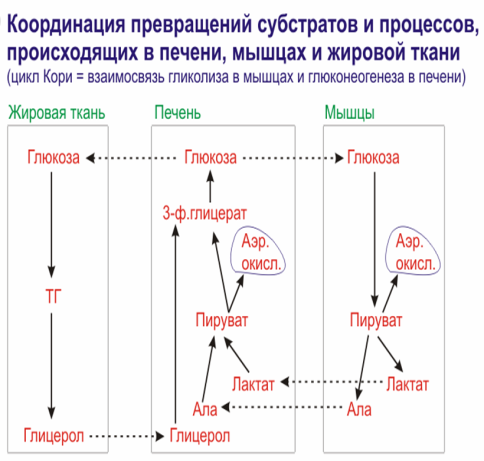
Физиологическая роль глюконеогенеза из лактата существенно иная. Молочная кислота не является конечным продуктом обмена, но ее образование — это тупиковый путь метаболизма: единственный способ использования молочной кислоты связан с ее превращением вновь в пируват при участии той же лактатдегидрогеназы:
СН3—СНОН—СООН + НАД+ -> СН3-СО-СООН + НАДН + Н*.
Из клеток, в которых происходит гликолиз, образующаяся молочная кислота поступает в кровь и улавливается в основном печенью, где и превращается в пируват. Пируват в печени частично окисляется, частично превращается в глюкозу — цикл Кори, или глюкозо-лактатпый цикл. Часть пирувата в мышцах путем трансаминирования превращается в аланин, который транспортируется в печень, и здесь снова образует пируват — глюкозоаланиповый цикл.
На каждую молекулу лактата при глюконеогенезе расходуется три молекулы АТФ (точнее, две — АТФ и одна — ГТФ) поскольку для образования глюкозы необходимо две молекулы лактата, суммарный процесс глюконеогенеза описывается так:
2Лактат + 6АТФ + 6Н20 -> Глюкоза + 6АДФ + 6Н3Р04,
Образовавшаяся глюкоза может вновь поступать в мышцы и там превращатется в молочную кислоту.
Регуляция гликолиза и глюконеогенеза.
Высокие концентрации АТФ и восстановленного НАДФ ингибируют гликолиз и увеличивают скорость глюконеогенеза.
Выысокие концентрации АДФ и АМФ стимулируют гликолиз и подавляют глюконеогенез.
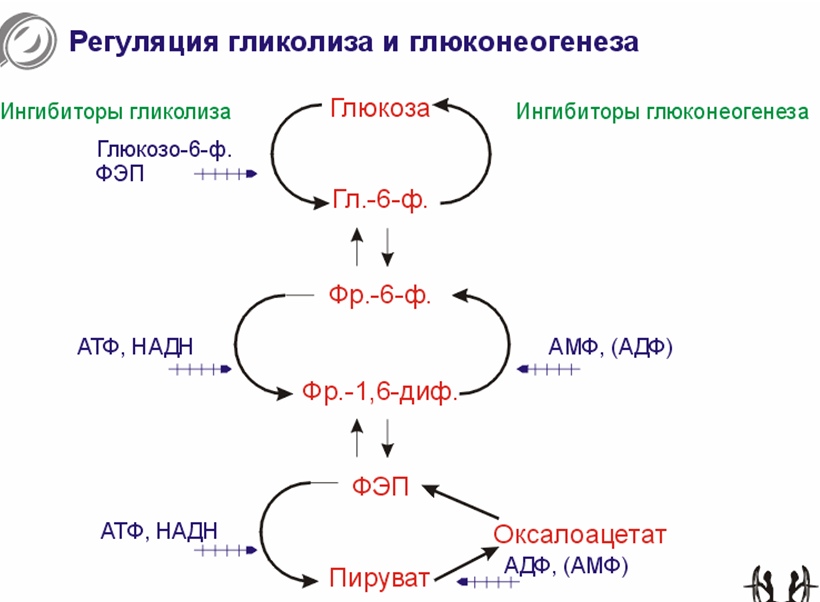
Общей чертой является интенсивнось от энергетического статуса клетки.
В мышцах при усиленной работе усиливается гликолиз, печень занимается процессами глюконеогенеза. Глюконеогенез в мышцах не идет. В печени и почках происходит глюконеогенез.
Вопрос 74. Биосинтез и мобилизация гликогена. Схема регуляции активности гликогенфосфорилазы и гликогенсинтазы.
Гликоген-высоковетвистый полимер с большой мол. массой. Это основное хранилище глюкозы в крови. Не создает осмотических проблем. Образуется, практически во всех тканях. Больше всего в гепатоците-4.8%. В печени хранится 50-100гр.
Биосинтез происходит в несколько стадий:

После удлинения цепи на 11-12 звеньев, 7 учясток переносится на внутренний учясток гликогена образуя связь (альфа)-1-6-появляется ветка. Катализирует фермент ветвления.