

59. Синтез и катаболизм гема. Значение конъюгирования продуктов метаболизма гема в печени.

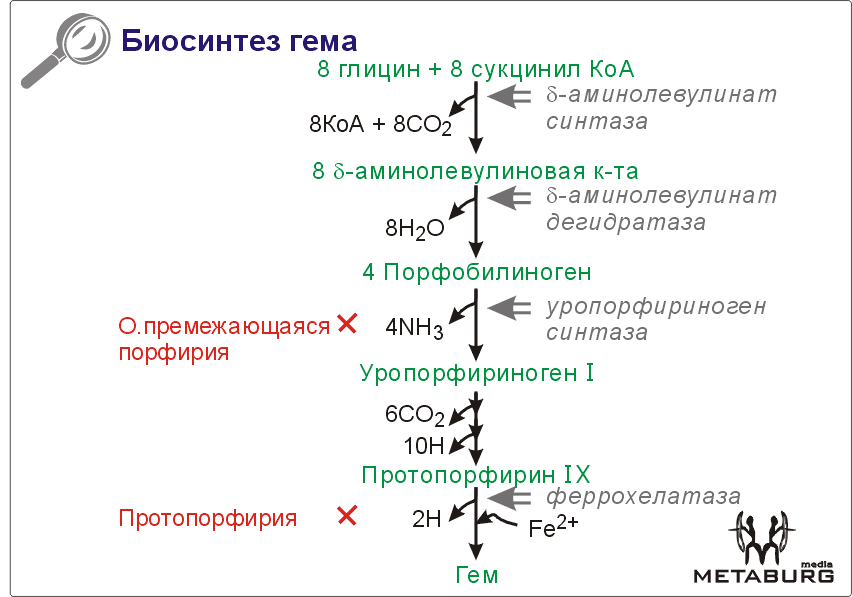
Гем, железосодержащее тетрагидропиррольное красящее вещество, является составной частью О2-связывающих белков и различных коферментов оксидоредуктаз .Почти на 85% биосинтез гема происходит в костном мозге и лишь небольшая часть — в печени. В синтезе гема участвуют митохондрии и цитоплазма.
Синтез тетрагидропиррольных колец начинается в митохондриях. Из сукцинил-КоА (на схеме наверху), промежуточного продукта цитратного цикла, конденсацией с глицином получается продукт, декарбоксилирование которого приводит к 5-аминолевулинату (ALA). Отвечающая за эту стадию 5-аминолевулинат-синтаза (ALA-синтаза)] является ключевым ферментом всего пути. Экспрессия синтеза ALA-синтазы тормозится гемом, т. е. конечным продуктом, и имеющимся ферментом. Это типичный случай торможения конечным продуктом, или ингибирования по типу обратной связи.
После синтеза 5-аминолевулинат переходит из митохондрий в цитоплазму, где две молекулы конденсируются в порфобилиноген, который уже содержит пиррольное кольцо.
На последующих стадиях образуется характерная для порфирина тетрапиррольная структура. Связывание четырех молекул порфобилиногена с отщеплением NH2-групп и образованием уропорфириногена III катализируется гидроксиметилбилан-синтазой . Для образования этого промежуточного продукта необходим второй фермент, уропорфириноген III-синтаза .Отсутствие этого фермента приводит к образованию «неправильного» изомера — уропорфириногена I.
Тетрапиррольная структура уропорфиринoгена III все еще существенно отличается от гема. Так, отсутствует центральный атом железа, а кольцо содержит только 8 вместо 11 двойных связей. Кроме того, кольца несут только заряженные боковые цепи R (4 ацетатных и 4 пропионатных остатков). Так как группы гема в белках функционируют в неполярном окружении, необходимо, чтобы полярные боковые цепи превратились в менее полярные. Вначале четыре ацетатных остатка (R1) декарбоксилируются с образованием метильных групп .Образующийся копропорфириноген III снова возвращается в митохондрии. Дальнейшие стадии катализируются ферментами, которые локализованы на/или внутри митохондриальной мембраны. Прежде всего под действием оксидазы две пропионатные группы (R2) превращаются в винильные Модификация боковых цепей заканчивается образованием протопорфириногена IX.
На следующей стадии за счет окисления в молекуле создается сопряженная π-электронная система, которая придает гему характерную красную окраску. При этом расходуется 6 восстановительных эквивалентов .В заключение с помощью специального фермента, феррохелатазы, в молекулу включается атом двухвалентного железа .Образованный таким образом гем или Fe-протопорфирин IX включается, например, в гемоглобин и миоглобин, где он связан нековалентно, или в цитохром С, с которым связывается ковалентно .
КАТАБОЛИЗМ ГЕМА: ОБРАЗОВАНИЕ ЖЕЛЧНЫХ ПИГМЕНТОВ
При физиологических условиях в организме взрослого человека разрушается 1--2108 эритроцитов в час. Таким образом, в течение суток у человека массой 70 кг обновляется приблизительно 6 г гемоглобина. При разрушении гемоглобина его белковая часть (глобин) может быть использована как таковая или после гидролиза в форме составляющих ее аминокислот; железо гема включается в общий пул железа и также снова используется. Вместе с тем свободная от железа порфириновая часть гема обязательно деградирует; это в основном происходит в ретикулоэндотелиальных клетках печени, селезенки и костного мозга. Катаболизм гема, освобожденного из любых гемовых белков, осуществляется в микросомальной фракции ретикулоэндотелиальных клеток сложной ферментной системой - гем-оксигеназой. К моменту поступления гема из гемовых белков в гем-оксигеназную систему железо обычно окисляется в ферри-форму (гем превращается в гемин); гемин может легко связываться с альбумином с образованием метгемальбумина. Гем-оксигеназная система индуцируется субстратом. Она локализована около микросомальной системы транспорта электронов. Гемин восстанавливается в ферро-форму с помощью NADPH; далее при участии NADPH кислород присоединяется к а-метенильному мостику между пиррольными кольцами I и II. Ферро-форма железа снова окисляется в ферри-форму. При последующем присоединении кислорода происходит освобождение ферри-иона, выделение молекулы оксида углерода (II) и образование в результате раскрытия тетрапиррольного кольца эквимолярного количества биливердина IX - . В этой реакции сам гем участвует в роли катализатора. Дальнейшей метаболизм билирубина в основном происходит в печени. Он складывается из трех процессов: 1) поглощение билирубина паренхимальными клетками печени; 2) конъюгация билирубина в гладком эндоплазматическом ретикулуме и 3) секреция билирубина из эндоплазматического ретикулума в желчь. Рассмотрим каждый из процессов в отдельности.
Поглощение билирубина печенью
Билирубин слаборастворим в плазме и воде; в плазме он специфически связывается с альбумином. Каждая молекула альбумина имеет, по-видимому, два центра связывания билирубина -- высоко- и низкоаффинный. В 100 мл плазмы может содержаться 25 мг билирубина, прочно связанного с альбумином по его высокоаффинному центру. «Избыточный» билирубин связывается с альбумином менее прочно; он легко отделяется от альбумина, диффундируя в ткани. Ряд соединений -- антибиотики и некоторые другие лекарственные вещества -- конкурируют с билирубином за высокоаффинный центр альбумина. Эти соединения могут вытеснять билирубин из комплекса с альбумином и проявляют значительное клиническое действие.
В печени происходит переход билирубина от альбумина на синусоидальную поверхность гепатоцитов при участии насыщаемой системы переноса, в функционировании которой участвует некий переносчик. Эта система облегченного транспорта имеет очень большую емкость и даже при патологических условиях не лимитирует скорость метаболизма билирубина. Поскольку система облегченного транспорта обеспечивает установление равновесия билирубина по обе стороны синусоидальной мембраны гепатоцита, поглощение билирубина зависит от его потребления в последующих метаболических процессах.

Конъюгация билирубина
В печени к билирубину присоединяются полярные группы и он переходит в водорастворимую форму, которая секретируется в желчь. Процесс, обеспечивающий повышение растворимости в воде (т. е. повышение полярности) билирубина, называется конъюгацией. Этот процесс, по крайней мере на начальных стадиях, протекает в гладком эндоплазматическом ретикулуме и осуществляется специальным набором ферментов. У млекопитающих билирубин секретируется в желчь преимущественно в форме билирубиндиглюкуронида
Секреция билирубина в желчь
Секреция конъюгированного билирубина в желчь идет против весьма высокого градиента концентрации и должна осуществляться с помощью механизма активного транспорта. Активный транспорт является, вероятно, скорость-лимитирующей стадией всего процесса метаболизма билирубина в печени. Транспорт конъюгированного билирубина из печени в желчь индуцируется теми же лекарствами, которые способны индуцировать конъюгацию билирубина. Таким образом, системы конъюгации билирубина и его вывода из гепатоцитов работают как единый функционально координируемый механизм.
При физиологических условиях практически весь секретируемый в желчь билирубин (свыше 97%) находится в конъюгированной форме. Только после светотерапии заметные количества неконъюгированного билирубина могут быть обнаружены в желчи.
В печени имеются многочисленные системы секреции в желчь природных и синтетических лекарственных соединений после их метаболизма. Некоторые из этих систем используются также билиру-биндиглюкуронидами.
МЕТАБОЛИЗМ БИЛИРУБИНА В КИШЕЧНИКЕ
После того как билирубин достигает области подвздошной и толстой кишок, глюкурониды гидролизуются специфическими бактериальными ферментами (р-глюкуронидазами); далее кишечная микрофлора восстанавливает пигмент с образованием группы бесцветных тетрапиррольных соединений, называемых уробилиногенами.
В подвздошной и толстой кишках небольшая часть уробилиногенов снова всасывается и попадает в печень, т.е. осуществляется внутрипеченочный уробилиногеновый цикл. При патологических состояниях, например при накоплении избыточных количеств желчных пигментов или при заболеваниях печени, нарушающих работу внутрипеченочного цикла, уробилиноген может экскретироваться с мочой.
В норме большая часть бесцветных уробилиногенов, образующихся в толстой кишке под действием кишечной микрофлоры, окисляется в уробилины (окрашенные соединения) и удаляется с фекалиями Наблюдаемое потемнение последних на воздухе является следствием окисления оставшихся уробилиногенов в уробилины.