

64. Механизм сопряжения окисления и фосфорилирования.
Как энергия(Е) электрон-дых цепи переходит в Е фосфатных связей АТФ? Существует несколько гипотез: 1) химич концепция – концеп промежут факторов фосрилирования – существуют промежуточные переносчики фосфорильных групп, сохранятся Е на обазование вязей. 2) конформационная – во время транспортировки ē энергия сохраняется внутри мемб митохондрии(МХ), к-ая «выбрасывает» эту Е при возвращении своей собственной конформации («пружина»). 3) хемоосмотическая (Митчелл, Скулачев, Овчинников, Уокер, Бойер) – внутр мемб МХ обладает высоким электрич сопротивлением и низкой проницаемостью для протонов и гидроксилов; окислительно-востановительные ē дых цепи располагаются поперек внутр мемб, по всей видимости изгибаясь и трижды ее прошнуровывая; предполагают, что в этой цепи имеются протонные транскиназы (протонные помпы); используя Е, выделяющуюся при переносе ē, эти помпы выкачивают протоны из матрикса в межмемб пространство; Е, к-ую отдают ē, частично рассеивается в виде тепла, запасается виде АТФ. Внутр мемб МХ уподобляется конденсатору: наруж сторона сопрягающей мемб получает «+» заряд, внутр сторона «-». На сопрягающей мемб одновременно с градиентом концентрации протонов возникает градиент электр потенциала. ΔµН+ - электро-хим градиент потенциала - конвентируемая форма Е, которая копится на внутр мемб МХ, хим Е окисляемых субстратов транспортир в электр Е и накапливается в форме электрич гадиента потенциала. Обратный поток протонов по градиенту их концентрации во внутрь матрикса осуществляется через спец канал АТФ-синтазы. Поток протонов и высвобождающаяся при этом Е служат движущей силой для синтеза АТФ.
Протоны от НАДН+Н+ идут на наружный листок внутр мемб МХ, а ē делают возвратную петлю с помощью FeS и переносится на ФМН. Для восстановления ФМН из матрикса потупают Н+.
Окончательный механизм транспорта Н+ не выяснен. Транслокация Н+ осуществляется 1,2,3 комплексами дых цепи (протонные транслоказы).
АТФ-синтаза катализирует фосфорилир АДФ до АТФ. Весь процесс окисления НАДН кислородом связан с переносом 10Н+ в межмемб пространство и прохождением через АТФсинтазный канал кажд 3Н+ обратно в матрикс, синтезируется 1мол АТФ.
Положения хемоосмотич теории: 1) создание электрон хим градиента Н+ на внутр мемб МХ является способом сопряжения потока Е, выделяющийся при транпортировке ē с запасанием ее в форме АТФ. 2) в то время как ē движутся вдоль дых цепи Н+ перемещаются из матрикса в межмемб пространство. 3) Н+ из межмемб пространства могут вернутся обратно в матрис только через спец канал, предоставленный АТФ-синтазой. Е, высвобождающаяся тогда, когда Н возвращаются по градиенту их концтр в матрикс обеспечивает фосфорилирование АДФ в АТФ.
65.Основной энергетический обмен и теплопродукция. Потоки важнейших метаболитов, поступающих в митохондрии и выходящих из них.
НАДН+Н+ + 1\2 О2 НАД+ + Н2О
320 кДж выделяется, 120 кДж запасается в виде АТФ, остальное рассеивается в виде тепла.
Сократительный термогенез – непроизвольное сокращение мышц (дрожание).
Основные источники теплоты – энергетич насосы, работающие в живой клетке; АТФ.
Основной обмен – состояние организма, когда теплопродукция является главным путем расходования энергии организма. До 10-12% Е приходится на синтетические процессы, 15% работа транспортных насосов, остальное – теплопродукция.
350 кДж\ч – интенсивность основного обмена.
П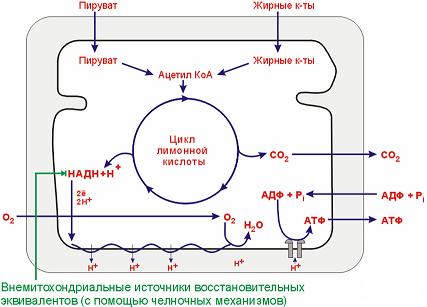 отоки
важнейших метаболитов, поступающих в
митохондрии и выходящих из них.
отоки
важнейших метаболитов, поступающих в
митохондрии и выходящих из них.
Челночные механизмы: α-глицерофосфатный; малатаспартатный.
Для окислительного метаболизма запасается дополнительное кол-во субстрата (гликогенов в печени и мышцах, липидов в жир ткани).