

Вопрос 60
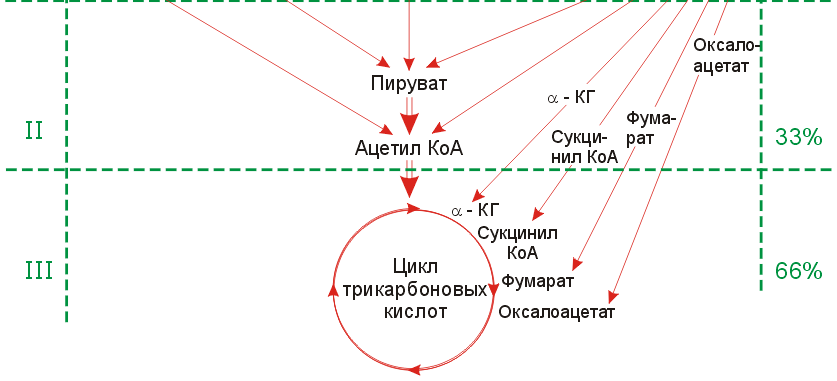
Общие и специфические пути катаболизма белков, жиров, углеводов. Схема трех основных стадий катаболизма ( по Кребсу ).
Стадия1(специфический путь катаболизма)

Стадия2 и 3(неспецифические пути катаболизма)
Вопрос 61
Окислительное декарбоксилирование пирувата. Пируватдегидрогеназный ферментативный комплекс.
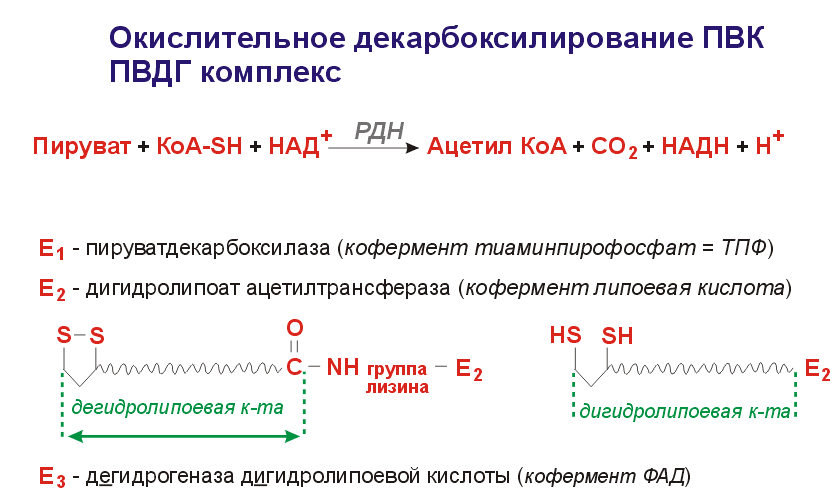
Окисление пирувата до ацетил-КоА происходит при участии ряда ферментов и коферментов, объединенных структурно в мультиферментную систему, получившую название «пируватдегидрогеназный комплекс».
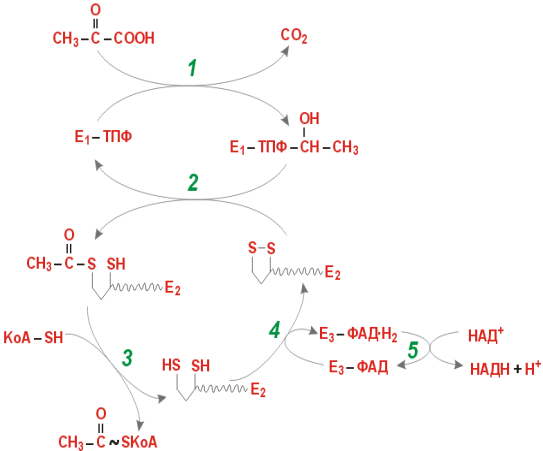
На I стадии этого процесса пируват теряет свою карбоксильную группу в результате взаимодействия с тиаминпирофосфатом (ТПФ) в составе активного центра фермента пируватдегидрогеназы (E1). На II стадии оксиэтильная группа комплекса E1–ТПФ–СНОН–СН3 окисляется с образованием ацетильной группы, которая одновременно переносится на амид липоевой кислоты (кофермент), связанной с ферментом дигидроли-поилацетилтрансферазой (Е2). Этот фермент катализирует III стадию – перенос ацетильной группы на коэнзим КоА (HS-KoA) с образованием конечного продукта ацетил-КоА, который является высокоэнергетическим (макроэргическим) соединением.
На IV стадии регенерируется окисленная форма липоамида из восстановленного комплекса дигидролипоамид–Е2. При участии фермента дигидролипоилдегидрогеназы (Е3) осуществляется перенос атомов водорода от восстановленных сульфгидрильных групп дигидролипоамида на ФАД, который выполняет роль простетической группы данного фермента и прочно с ним связан. На V стадии восстановленный ФАДН2 дигидро-липоилдегидрогеназы передает водород на кофермент НАД с образованием НАДН + Н+.
Процесс окислительного декарбоксилирования пирувата происходит в матриксе митохондрий. В нем принимают участие (в составе сложного мультиферментного комплекса) 3 фермента (пируватдегидрогеназа, ди-гидролипоилацетилтрансфераза, дигидролипоилдегидрогеназа) и 5 кофер-ментов (ТПФ, амид липоевой кислоты, коэнзим А, ФАД и НАД), из которых три относительно прочно связаны с ферментами (ТПФ-E1, ли-поамид-Е2 и ФАД-Е3), а два – легко диссоциируют (HS-KoA и НАД).
Регуляция ПВДГ комплекса.

Вопрос 62
Цикл лимонной кислоты; локализация, регуляция, функции.
Цикл лимонной кислоты

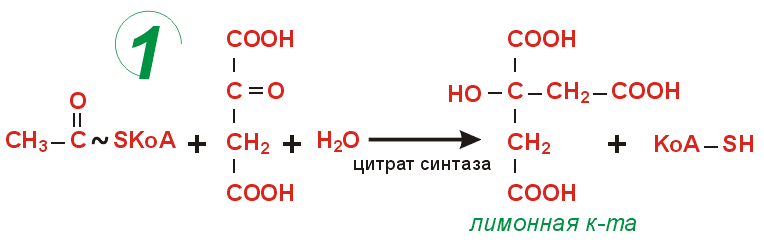
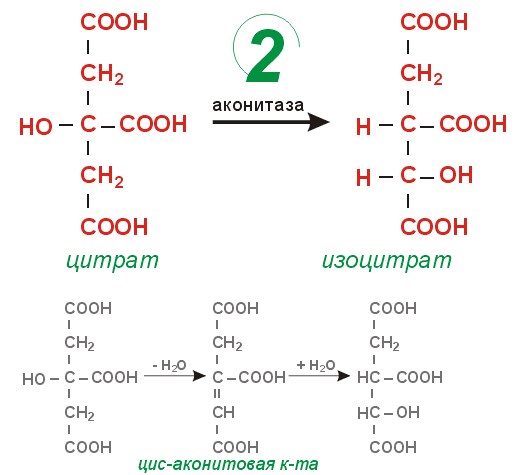

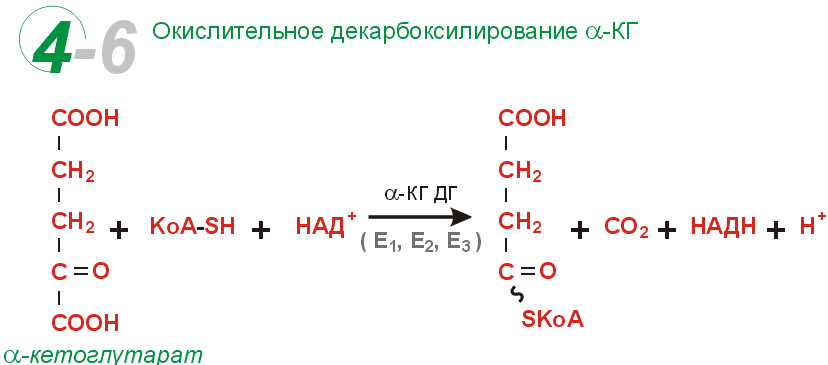

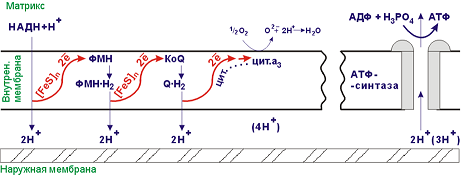

Цикл Кребса сложный цикл реакций, где в качестве катализаторов выступают ферменты; эти реакции проходят в клетках всех животных и заключаются в разложении ацетата в присутствии кислорода с выделением энергии в виде АТФ (по цепи передачи электронов) и углекислого газа. Этот цикл является заключительным этапом окисления углеводов, жиров и белков; некоторые промежуточные продукты этого цикла используются в процессе синтеза аминокислот в организме.цикл лимонно-кислый
Регуляция и функция цикла лимонной кислоты
Л![]() имонная
кислота, являясь главным промежуточным
продуктом метаболического цикла
трикарбоновых кислот, играет важную
роль в системе биохимических реакций
клеточного дыхания множества организмов.
Едва ли можно сомневаться в том, что в
большинстве тканей, в которых основная
функция цикла лимонной кислоты -
обеспечение энергией, дыхательный
контроль, осуществляемый при
функционировании дыхательной цепи и
окислительного фосфорилирования ,
является определяющим фактором при
регуляции активности рассматриваемого
цикла. Активность этого цикла
непосредственно связана с поступлением
окисленных кофакторов дегидрогеназ
(например, NAD ), которое в свою очередь
зависит от доступности ADP и, в конечном
счете, от скорости потребления ATP .
Свойства ряда ферментов этого цикла
указывают на то, что кроме общей регуляции
существует также регуляция на уровне
самого цикла. В клетках головного мозга
, в которых ацетил-CoA образуется в основном
из углеводов , регуляция цикла лимонной
кислоты может происходить на стадии,
катализируемой пируватдегидрогеназой
. В самом цикле регуляция может
осуществляться путем аллостерического
ингибирования цитратсинтазы при действии
ATP или ацил-CoA-производных длинноцепочечных
жирных кислот . Митохондриальная
NAD-зависимая изоцитратдегидрогеназа
аллостерически активируется ADP и
ингибируется ATP и NADH .
Альфа-кетоглутаратдегидрогеназный
комплекс регулируется, по-видимому,
аналогично пируватдегидрогеназе.
Сукцинатдегидрогеназа ингибируется
оксалоацетат ом, а образование
оксалоацетата в малатдегидрогеназной
реакции зависит от соотношения
[NADH]/[NAD+] . Поскольку величина Km цитратсинтазы
для оксалоацетата такого же порядка,
что и величина внутримитохондриальной
концентрации оксалоацетата, концентрация
последнего, по-видимому, играет
определенную роль в регуляции скорости
образования цитрат а. Какие из
вышеперечисленных механизмов регуляции
в самом цикле функционируют in vivo, пока
еще не ясно.
имонная
кислота, являясь главным промежуточным
продуктом метаболического цикла
трикарбоновых кислот, играет важную
роль в системе биохимических реакций
клеточного дыхания множества организмов.
Едва ли можно сомневаться в том, что в
большинстве тканей, в которых основная
функция цикла лимонной кислоты -
обеспечение энергией, дыхательный
контроль, осуществляемый при
функционировании дыхательной цепи и
окислительного фосфорилирования ,
является определяющим фактором при
регуляции активности рассматриваемого
цикла. Активность этого цикла
непосредственно связана с поступлением
окисленных кофакторов дегидрогеназ
(например, NAD ), которое в свою очередь
зависит от доступности ADP и, в конечном
счете, от скорости потребления ATP .
Свойства ряда ферментов этого цикла
указывают на то, что кроме общей регуляции
существует также регуляция на уровне
самого цикла. В клетках головного мозга
, в которых ацетил-CoA образуется в основном
из углеводов , регуляция цикла лимонной
кислоты может происходить на стадии,
катализируемой пируватдегидрогеназой
. В самом цикле регуляция может
осуществляться путем аллостерического
ингибирования цитратсинтазы при действии
ATP или ацил-CoA-производных длинноцепочечных
жирных кислот . Митохондриальная
NAD-зависимая изоцитратдегидрогеназа
аллостерически активируется ADP и
ингибируется ATP и NADH .
Альфа-кетоглутаратдегидрогеназный
комплекс регулируется, по-видимому,
аналогично пируватдегидрогеназе.
Сукцинатдегидрогеназа ингибируется
оксалоацетат ом, а образование
оксалоацетата в малатдегидрогеназной
реакции зависит от соотношения
[NADH]/[NAD+] . Поскольку величина Km цитратсинтазы
для оксалоацетата такого же порядка,
что и величина внутримитохондриальной
концентрации оксалоацетата, концентрация
последнего, по-видимому, играет
определенную роль в регуляции скорости
образования цитрат а. Какие из
вышеперечисленных механизмов регуляции
в самом цикле функционируют in vivo, пока
еще не ясно.