

40. Репликация днк у эукариот. Свойства днк-полимераз эукариот. Репликация днк и клеточный цикл.
Особенности репликации ДНК у эукариот. Репликация ДНК у эукариот, по существу аналогичная репликации ДНК у прокариот, имеет ряд особенностей. Например, вместо одной точки репликации в ДНК эукариот имеются специфические точки «начала», так называемые автономно реплицирующие последовательности (около 300 нуклеотидных пар); в дрожжевой клетке таких элементов около 400. Кроме того, скорость движения репликационной вилки у эукариот (примерно 50 нуклеотидов в секунду) почти в 10 раз ниже, чем у E. coli. Для репликации ДНК генома человека из одной-единственной точки с подобной скоростью потребовалось бы более 500 ч; вместо этого репликация генома человека происходит в обоих направлениях и одновременно из множества точек (множество «начал» репликации), вовлекая от 30000 до 330000 пар оснований. Репликация продолжается до тех пор, пока не будут синтезированы две дочерние молекулы ДНК, в каждой из которых содержится одна родительская цепь (см. рис. 13.4). Таким образом, множественность точек «начала» репликации ДНК, вероятнее всего, является общим правилом для всех клеток эукариот.
Рис. 13.2. Полуконсервативная репликация ДНК in vitro. Каждая из двух цепей родительской ДНК служит матрицей для синтеза дочерних молекул ДНК. 1 - родительская молекула; 2 -дочерние молекулы (первая генерация); 3 - дочерние молекулы (вторая генерация).

Рис. 13.3. Основные этапы репликации ДНК (схема).

Рис. 13.4. Схематическое изображение непрерывного и прерывистого синтеза цепей ДНК при репликации.
Как было указано, инициация биосинтеза дочерних цепей ДНК требует предварительного синтеза на матрице ДНК необычного затравочного олигорибонуклеотида, названного праймером, со свободной гидроксильной группой у С-3' рибозы. Этот короткий олигорибонуклеотид синтезируется комплементарно на матрице ДНК при участии особого фермента – прай-мазы, наделенной РНК-полимеразной активностью.
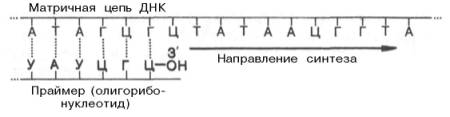
Предполагают, что именно с этой точки концевого 3'-гидроксила рибозы праймера начинается истинный синтез лидирующей дочерней цепи ДНК, комплементарной родительской. Синтез начинается с реакции между 3'-ОН-группой концевого рибонуклеотида праймера и α-фосфатной группой первого дезоксирибонуклеотидтрифосфата в строгом соответствии с комплементарностью родительской цепи ДНК, при этом освобождается пирофосфат. В дальнейшем этот фрагмент РНК, комплементарно присоединенный к новообразованной цепи ДНК, разрушается под действием ДНК-полимеразы I, и возникшая брешь застраивается олигодезоксирибо-нуклеотидом при помощи той же ДНК-полимеразы I. Вполне допустимо предположение, что синтез праймера из олигорибонуклеотида имеет глубокий биологический смысл, поскольку в этом случае могут устраняться ошибки, неизбежно возникающие при инициации репликации ДНК.
Этапы биосинтеза ДНК. Предложен ряд моделей механизма биосинтеза ДНК с участием указанных ранее ферментов и белковых факторов, однако детали некоторых этапов этого синтеза еще не выяснены. Основываясь главным образом на данных, полученных в опытах in vitro, предполагают, что условно механизм синтеза ДНК у Е. coli может быть подразделен на три этапа; инициацию, т.е. начало, элонгацию, т.е. продолжение, и терми-нацию, т.е. завершение (прекращение) синтеза. Каждый из этих этапов требует участия специфических ферментов и белковых факторов.
Этап I – инициация биосинтеза ДНК – является началом синтеза дочерних нуклеотидных цепей; в инициации участвует минимум восемь хорошо изученных и разных ферментов и белков. Первая фаза – это, как указано ранее, ферментативный биосинтез на матрице ДНК необычного затравочного олигорибонуклеотида (праймера) со свободной гидроксиль-ной группой у С-3' рибозы. При инициации к цепям ДНК последовательно присоединяются ДНК-раскручивающие и ДНК-связывающие белки, а затем комплексы ДНК-полимераз и праймаз (см. рис. 13.3). Инициация представляется единственной стадией репликации ДНК, которая весьма тонко и точно регулируется, однако детальные механизмы ее до сих пор не раскрыты и в настоящее время интенсивно исследуются.
Этап II – элонгация синтеза ДНК – включает два кажущихся одинаковыми, но резко различающихся по механизму синтеза лидирующей и отстающей цепей на обеих материнских цепях ДНК. Синтез лидирующей цепи начинается с синтеза праймера (при участии праймазы) у точки начала репликации, затем к праймеру присоединяются дезоксирибонуклеотиды под действием ДНК-полимеразы III; далее синтез протекает непрерывно, следуя шагу репликационной вилки. Синтез отстающей цепи, напротив, протекает в направлении, обратном движению репликационной вилки и начинается фрагментарно. Фрагменты всякий раз синтезируются раздельно, начиная с синтеза праймера, который может переноситься с готового фрагмента при помощи одного из белковых факторов репликации в точку старта биосинтеза последующего фрагмента противоположно направлению синтеза фрагментов. Элонгация завершается отделением олигорибонуклеотидных праймеров, объединением отдельных фрагментов ДНК при помощи ДНК-лигаз и формированием дочерней цепи ДНК. Нельзя исключить, однако, возможности сопряженного и согласованного механизма синтеза лидирующей и отстающей цепей ДНК при участии полимераз и всего комплекса праймасом.
Этап III – терминация синтеза ДНК – наступает, скорее всего, когда исчерпана ДНК-матрица и трансферазные реакции прекращаются. Точность репликации ДНК чрезвычайно высока, возможна одна ошибка на 1010 трансферазных реакций, однако подобная ошибка обычно легко исправляется за счет процессов репарации.
Синтез ДНК на матрице РНК. Выдающимся достижением биохимии нуклеиновых кислот является открытие в составе онковирусов (вирус Раушера и саркомы Рауса) фермента обратной транскриптазы, или ревертазы (РНК-зависимая ДНК-полимераза), катализирующего биосинтез молекулы ДНК на матрице РНК. Накоплены данные о том, что многие РНК-содержащие онкогенные вирусы, получившие наименование онкорнавирусов, содержат ревертазу в составе покровных белков. Фермент открыт также во многих клетках прокариотов и эукариотов, в частности в лейкозных клетках, пролиферирующих тканях, включая эмбриональные ткани. Ревертаза онкорнавирусов содержит ионы Zn2+и активируется катионами Мn2+ и Mg2+ . Предполагают, что синтез ДНК на матрице РНК происходит в 3 этапа. На I этапе фермент ревертаза синтезирует на матрице вирусной РНК комплементарную цепь ДНК, что приводит к формированию гибридной молекулы. Второй этап – разрушение исходной вирусной РНК из комплекса гибридной молекулы под действием РНКазы. Наконец, на III этапе на матрице цепи ДНК комплементарно синтезируются новые цепи ДНК. Ревертазной активностью обладают и ДНК-полимеразы: например, фермент из Е. coli способен катализировать синтез ДНК на матрице рРНК.
Открытие обратной транскриптазы имеет большое значение не только для выяснения закономерностей процесса малигнизации, но и для всей науки о живом, поскольку указывает на возможность передачи наследственной информации от РНК на ДНК, не подчиняясь основному постулату (поток информации идет только в одном направлении):
ДНК –> РНК –> Белок.
В настоящее время можно дополнить эту основную схему передачи генетической информации в живой клетке и представить ее в более полной форме:
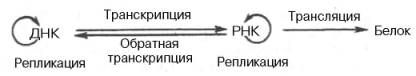
На схеме стрелки вокруг ДНК и РНК указывают на возможность молекул копировать самих себя в живых системах при участии соответствующих ферментов. Как знать, не станем ли мы свидетелями открытия принципиальной возможности поворота стрелки и на следующей стадии – от белка на РНК, что могло происходить на Земле при зарождении первичных живых существ?