

Вопрос 24. Зависимость скорости ферментативной реакции от рН, температуры, концентрации фермента, субстрата.
 Линейная
зависимость
Линейная
зависимость
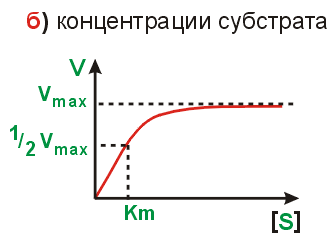
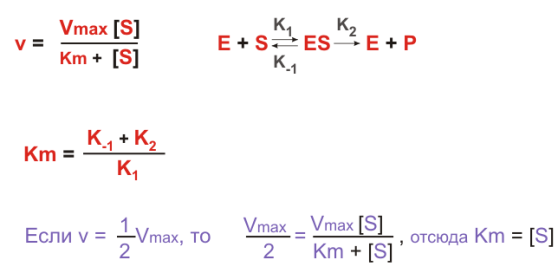
Vmax – активные центры всех молекул
Увеличение [S] накапливающегося в рез-те реакции ↓ V реакции
При небольших конц. [S] – реакции 1-го порядка.(v не зависит от конц [S])
Математическое описание зависимости V и S. Бриттс Холдейн
Для односубстратных реакций в условиях стационарной кинетики
Накапл. [S]
Конц. Продуктов ph мала (нет ингибиторов)
v→s ФСК=v его распада
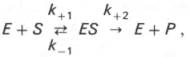
Кm представляет собой константу Михаэлиса, являющуюся экспериментально определяемой величиной. Она может быть представлена следующим уравнением:

k-1 – скорость распада
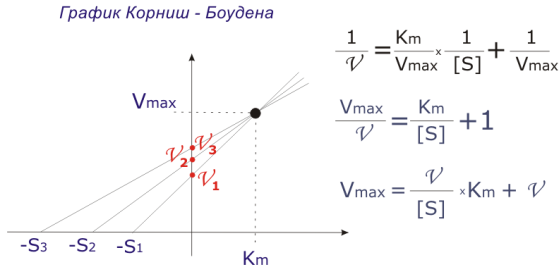
по константе михаэлиса судят о степени сродства субстрата и фермента. Чем ниже Km значит тем быстрее идёт ферм.р-ция. Графический способ опр. Графический способ определения Km.





 ph-оптимум-
значение ph
при котором реакция протекает наиболее
быстро. Для большинства E
он лежит в пределах от 6 до 8
ph-оптимум-
значение ph
при котором реакция протекает наиболее
быстро. Для большинства E
он лежит в пределах от 6 до 8

28.Регуляция активности ферментов. Ключевые ферменты-ферменты, запускающие метаболический путь, а также ферменты, лимитирующие скорость его протекания. Лимитирующая реакция – реакция с самой высокой энергией активации. Факторы регуляции скорости протекания метаболического пути:1)общие условия регулируют активность Ф. – концентрация Ф. и субстрата (S), значение рН,t0,времени;2)Аллостерическая регуляция - специальные, специфические активаторы и ингибиторы являются эффекторами (фактор активации – работа АЦ и S;фактор ингибирования – несоответствие АЦ и S). Эффектор может взаимодействовать непосредственно с каталитической субъединицей, и с регуляторной субъединицей, которая дает команду к формированию каталитической субъединице. Объединение субъединиц в олигомерный комплекс усиливает чувствительность Ф в ответ на малое колебание концентрации эффекторов. Олигомерность приводит к явлению кооперативности. Присоединение облегчает последующее взаимодействие субъед. и S. Олигомерность ускоряет или тормозит активность Ф в ответ на незначительные колебания концентрации S. 3)Активация предшественников (форактивация): 1 метаболит активирует Ф, катализирующий последующую стадию (необязательно ближайшую).4)ретроингибирование – торможение по принципу обратной связи, адекватная быстрая регуляция прямого метаболического пути.5)хим.модификация белков – Ф: к белку-Ф ковалентно присоединятся хим.функц.группа, что активирует или ингибирует деятельность Ф. Ф находится в активном состоянии несколько минут, а затем модифицированная группа удаляется с помощью лиаз.6)Хроническая регуляция – подавление синтеза ненужного в данный момент Ф, активация синтеза нужного; контроль на уровне генома: транскрипция/трансляция.7)регуляция активности Ф через гормоны: гормон-первичный сигнализатор→ рецептор→ регуляция активности.8) компартментализация: в клетке Ф и S могут быть разделены мембраной. Любой фактор, оказывающий влияние на проницаемость мембраны, будет являться регулятором активности ФК. Удаление продукта реакции (пространственное разообщение) также влияет на скорость. В случае разделения мембраной продукта и Ф, реакция протекает под ингибирование др. влияний.9)активация Ф путем протеолиза – для формирования АЦ
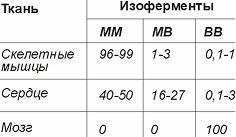
29. Изоферменты и целесообразность определения изоферментного спектра в медицине. Изоферм – разные молекуляр формы одного и того же ферм, кодирующиеся родственными генами. Отличаются рядом структурных, катализирующих и метаболич св-в: разные ферм имеют разные рН-оптимум, отличаются степенью сродства к субстрату и кофакторам, чувствит к активаторам и ингибиторам, скоростью синтеза и распада. Благодаря набору изоферм клетка обладает способностью к тонкой адаптации. Тканевые изоферм отвечают за приспосабливание ткани и оптимальное протекание р-ии при имеющихся условиях. 1) Изоферм могут быть гибридами двух видов полипепт цепей: креатинкиназа, ЛДГ, изоферм имеющие более 2-х полипепт цепочек. 2) аллелозины – генетич вариант одного и того же ферм, встречается у организмов – глюкозо-6-фосфат-ДГ. 3) генетич независимые изоферм, кодируются разными генами, может быть разная внутриклеточная организация (малатДГ,аминотрансфераза).

30.
Мультиферментные системы. Локализация
Ф:
мультиФ системы – надмолекулярный
комплекс из нескольких Ф, которые
работают сообща. Продукт 1 Ф→S
для 2![]() продукты
2 Ф
S
для 3. 1
тип:
Ф свободно расположены в цитоплазме –
все Ф гликолиза, Ф фосфоглюконатного
пути. 2
тип:
Ф объединены в комплекс с
определ.пространственной организацией.
S,
попав в комплекс, не покидает его пока
не образуется конечный продукт:
синтетазный комплекс жирных кислот,
пируватдегидрогеназный комплекс.3
тип:
отдельные Ф мультиФ комплекса присоединены
к опр.клеточным структурам: отд. Ф
дых.цепи связаны с внутр. Мембраной МХ,
а др. Ф – нет; отдельные Ф синтеза белка
– в рибосомах, а др. – где-то рядом.
Локализация Ф: внутри
клетки
- в составе цитоплазмы в опр.органеллах,
встроены в мембрану. Могут быть отдельные
отсеки, различающиеся по метаболитическим
процессам. Все Ф гликолиза – в цитоплазме,
Ф ЦТК, β-окисления жирных кислот – в
матриксе МХ. Органоспецифические Ф –
в одном или нескольких органах.
продукты
2 Ф
S
для 3. 1
тип:
Ф свободно расположены в цитоплазме –
все Ф гликолиза, Ф фосфоглюконатного
пути. 2
тип:
Ф объединены в комплекс с
определ.пространственной организацией.
S,
попав в комплекс, не покидает его пока
не образуется конечный продукт:
синтетазный комплекс жирных кислот,
пируватдегидрогеназный комплекс.3
тип:
отдельные Ф мультиФ комплекса присоединены
к опр.клеточным структурам: отд. Ф
дых.цепи связаны с внутр. Мембраной МХ,
а др. Ф – нет; отдельные Ф синтеза белка
– в рибосомах, а др. – где-то рядом.
Локализация Ф: внутри
клетки
- в составе цитоплазмы в опр.органеллах,
встроены в мембрану. Могут быть отдельные
отсеки, различающиеся по метаболитическим
процессам. Все Ф гликолиза – в цитоплазме,
Ф ЦТК, β-окисления жирных кислот – в
матриксе МХ. Органоспецифические Ф –
в одном или нескольких органах.