

График рецепторного потенциала (рп)
1) медленная деполяризация, 2) медленная реполяризация
Чем интенсивнее стимуляция, тем выше амплитуда РП, больше сила местных ионных токов и выше частота серии потенциалов действия (ПД) в афференте. При этом между амплитудой РП и частотой импульсной активности афферентного нейрона существует логарифмическая или степенная зависимость. Поэтому между силой раздражителя, действующего на сенсорные рецепторы, и выраженностью ощущения, возникающего в коре больших полушарий головного мозга, также отмечается логарифмическая или степенная, а не прямая зависимость.
Вторичночувствующие рецепторы представлены специализированными рецепторными клетками (фоторецепторными, фонорецепторными, вкусовыми и вестибулярными сенсорными клетками), которые образуют синаптические контакты с периферическими отростками первичных афферентных нейронов.
Вторичночувствующие рецепторы
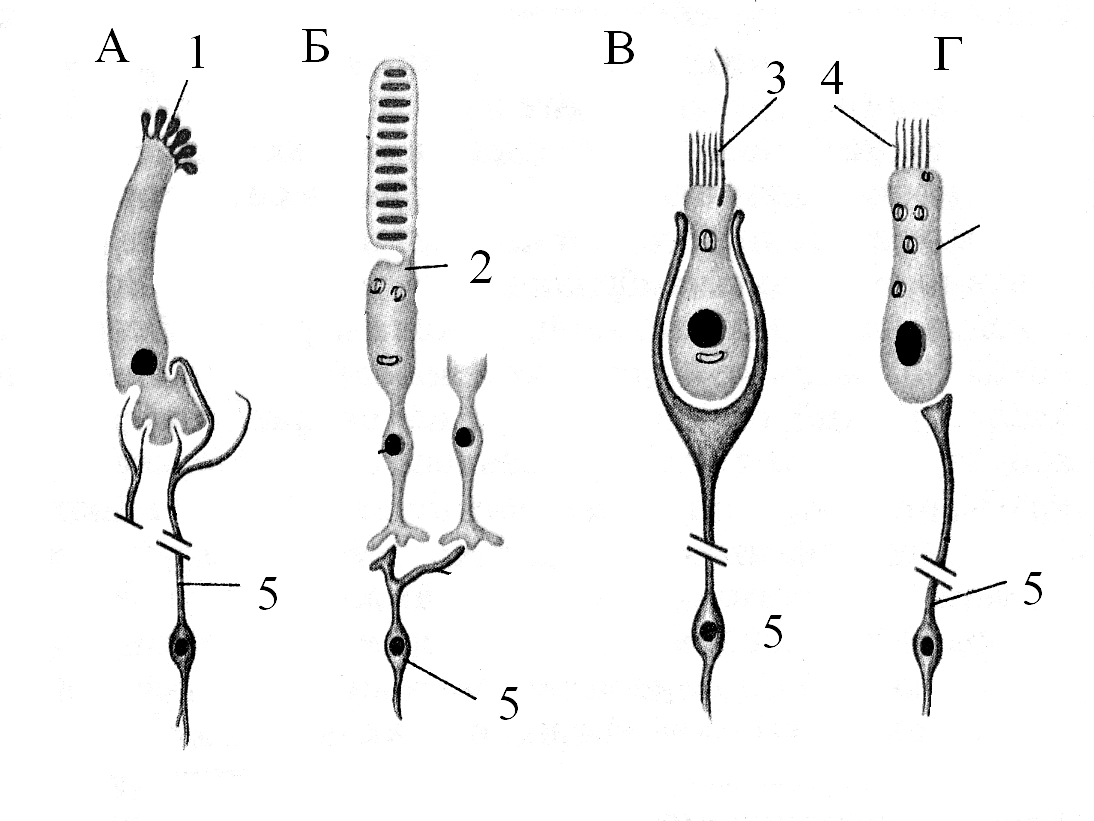
А – вкусовой рецептор, Б – фоторецептор, В – вестибулярный рецептор, Г – слуховой рецептор, 1 – вкусовая сенсорная клетка, 2 – фотосенсорная клетка, 3 – вестибулярная сенсорная клетка, 4 - фоносенсорная клетка, 5 – первичные афференты.
Механизм трансдукции сенсорного сигнала во вторичночувствующем рецепторе
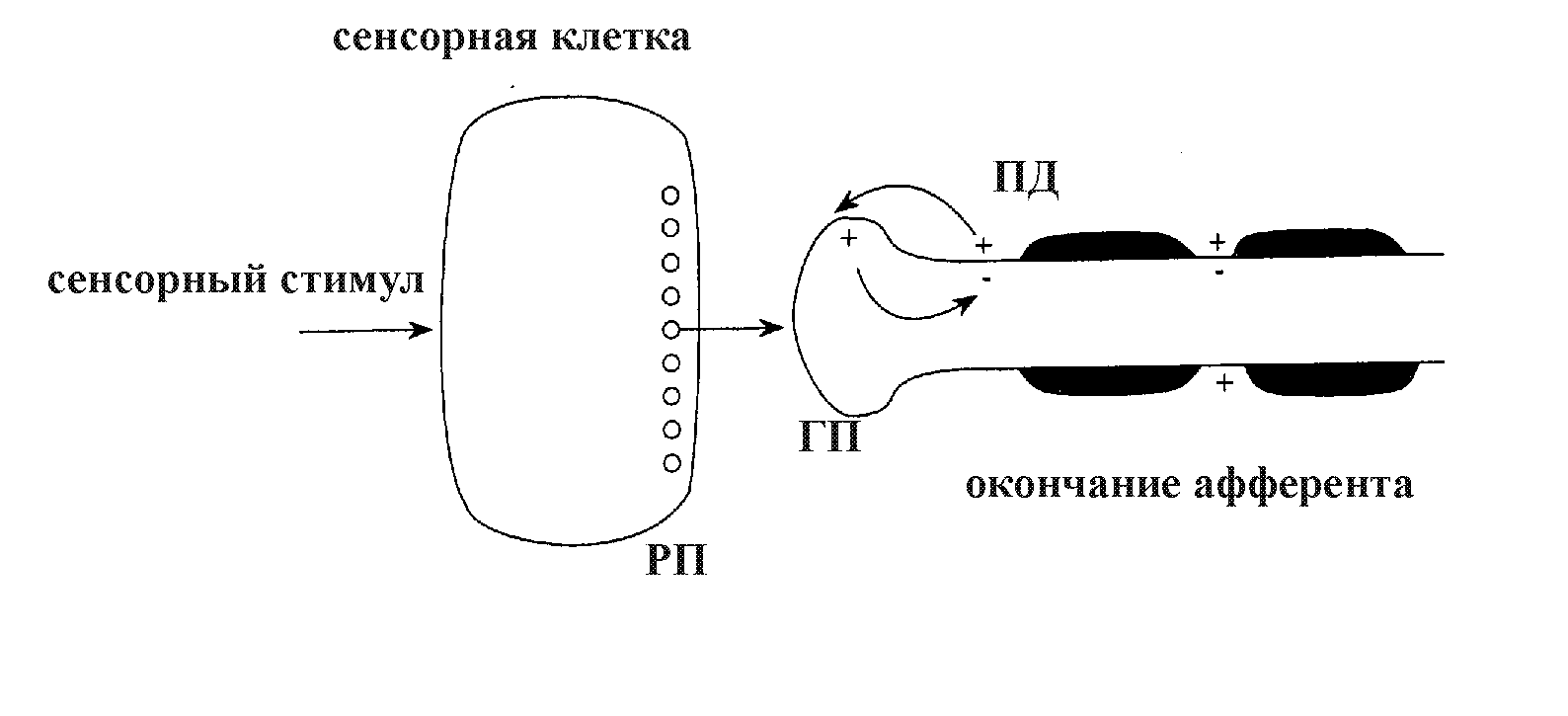
РП – рецепторный потенциал, ГП – генераторный потенциал, ПД – потенциал действия.
Большинство рецепторных клеток реагируют на действие раздражителя частичной деполяризацией - генерацией рецепторного потенциала. В результате деполяризации из рецепторной клетки выделяется возбуждающий медиатор, который диффундирует через синаптическую щель к постсинаптической мембране, расположенной на окончании периферического отростка чувствительного афферентного нейрона. По принципу комплементарности медиатор взаимодействует со специфическими белковыми молекулами-рецепторами постсинаптической мембраны рецепторно-афферентного синапса, что вызывает активацию ее хемовозбудимых натриевых каналов. Вследствие этого увеличивается проницаемость окончания афферента для ионов Na+ и происходит его частичная деполяризация, которая называется генераторным потенциалом. Затем благодаря циркуляции местных ионных токов между частично деполяризованным окончанием афферента и первым перехватом Ранвье чувствительный нейрон генерирует серию афферентных ПД.
Сенсорные рецепторы обладают тремя общими свойствами: высокой специфичностью, способностью к адаптации и функциональной мобильностью.
Специфичность - это способность рецептора воспринимать только адекватные раздражители.
Адекватными называют раздражители, к восприятию которых организм специально приспособился в процессе эволюции. Такие сенсорные стимулы вызывают возбуждение даже при очень малой интенсивности.
Для фоторецепторов адекватным раздражителем являются световые фотоны, для слуховых рецепторов – акустические волны, для обонятельных рецепторов – летучие, а для вкусовых – растворимые химические вещества.
Если сенсорный рецептор не приспособлен к восприятию какого-либо раздражителя, то данный фактор является для него неадекватным. Неадекватными называют раздражители, которые не являются в естественных условиях фактором возбуждения рецепторов.
Фонорецепторы и волосковые чувствительные клетки вестибулярного аппарата могут возбуждаться колебаниями с амплитудой, сопоставимой с диаметром атома водорода, а обонятельный рецептор – 1-2 молекулами пахучего вещества. На одиночные молекулы растворенного вещества реагирует и вкусовая сенсорная клетка.
Для возникновения ощущения света достаточно 1-2 фотонов с мощностью всего 10-17 - 10-18 Вт, т.к. они являются адекватным раздражителем для органа зрения. В то же время, ощущение света – механический фосфен, может возникнуть и при резком механическом воздействии на орган зрения («искры из глаз»), хотя никакого света на самом деле не было. Однако для этого необходимо приложить значительное усилие с мощностью более 10-4 Вт, т.к. фоторецепторы глаза не приспособлены специально для обнаружения такого раздражителя такой природы. Следовательно, разница между адекватным световым и неадекватным механическим пороговыми раздражителями для фоторецепторов глаза человека достигает 14 порядков.
Функциональная мобильность - способность сенсорной системы изменять количество активно функционирующих рецепторов, а также регулировать величину порога возбудимости каждого из них.
Механизм функциональной мобильности обусловлен возбуждающим или тормозным эфферентным нисходящим влиянием на сенсорные рецепторы и афференты со стороны коры и подкорковых структур головного мозга. Благодаря функциональной мобильности создаются благоприятные условия для восприятия наиболее значимых в данный момент времени сенсорных раздражителей. Так, у голодного человека возрастает число активно функционирующих вкусовых рецепторов и повышается чувствительность каждого из них. Поэтому во время приема пищи возникают яркие и сильные вкусовые ощущения. Напротив, у сытого человека выраженность и интенсивность вкусовых ощущений снижается.
При длительном воздействии любого раздражителя, вызванное им ощущение постепенно уменьшается, а иногда и полностью исчезает. Так, войдя в помещение, в котором проводились покрасочные работы, человек через некоторое время перестает ощущать столь резкий вначале запах краски. Точно также мы не замечаем непрерывного давления на кожу привычной одежды или яркого света, который вначале ослепляет нас. Это явление получило название сенсорная адаптация.
Сенсорная адаптация – это уменьшение чувствительности рецепторов, обусловленное снижением амплитуды рецепторного потенциала и частоты разрядной деятельности афферентного нейрона при длительном действии раздражителя.
Снижение амплитуды рецепторного потенциала и частоты импульсной биоэлектрической активности афферента связано со свойствами дорецепторного вспомогательного аппарата, а также изменениями процессов рецепции и трансдукции сенсорного сигнала в рецепторах.
Дорецепторное звено анализатора – специальный анатомический аппарат, позволяющий регулировать поступление энергии стимула к рецепторам. При длительном действии раздражителя дорецепторный аппарат ограничивает поступление энергии сенсорного стимула к рецепторам. Так, в условиях яркой освещенности происходит сужение зрачка, а постоянный звуковой шум вызывает сокращение мышц, уменьшающих амплитуду колебания барабанной перепонки.
Снижение уровня готовности к рецепции и трансдукции сенсорного сигнала в первичных рецепторах обусловлено:
падением чувствительности рецепторной мембраны, связанным с инактивацией встроенных в нее рецептирующих белковых молекул,
угнетением генеративных свойств - способности генерировать биоэлектрические потенциалы.
Снижение способности сенсорного рецептора генерировать биопотенциалы, обусловлено ионной адаптацией – инактивацией натриевых и активацией калиевых мембранных каналов, что приводит к снижению амплитуды РП и частоты разрядной деятельности афферента.
Во вторичночувствующих рецепторах все это усугубляется утомлением в синаптических контактах между специализированными сенсорными клетками и окончаниями афферентных нейронов:
истощением запасов медиатора в рецепторной клетке,
уменьшением чувствительности рецепторных белковых молекул постсинаптической мембраны,
недостаточностью ресинтеза медиатора.
В зависимости от скорости адаптации выделяют три вида сенсорных рецепторов: быстроадаптирующиеся, неадаптирующиеся, медленноадаптирующиеся.
Быстроадаптирующиеся рецепторы (например, тактильные и температурные рецепторы кожи) характеризуются быстрым снижением амплитуды РП и частоты разрядной деятельности афферента при длительном действии раздражителя. Неадаптирующиеся рецепторы (например, барорецепторы и хеморецепторы сосудов) характеризуются практическим отсутствием изменений РП и разрядной деятельности афферента при длительном действии раздражителя. Медленноадаптирующиеся рецепторы (например, тонические проприорецепторы интрафузальных мышечных волокон, сухожилий, связок) характеризуются постепенным снижением величины РП и частоты разрядной деятельности афферента при длительном действии раздражителя.
Адаптация происходит не только в рецепторах, но и в проводниковом, а также центральном отделах анализатора. В проводниковом отделе анализатора адаптации развивается за счет снижения чувствительности к медиатору постсинаптических мембран в спинальных, стволовых и подкорковых нервных центрах, участвующих в обработке сенсорных сигналов. В корковом отделе адаптация связана с выключением из сферы сознания и анализа несущественной для организма информации.
Проводниковый отдел анализатора представлен проводящими путями, а также спинальными, стволовыми и подкорковыми сенсорными центрами. Каждая область мозга, в которой находится сенсорный центр и осуществляется переключение проводящих путей, образует уровень сенсорной системы. Высший подкорковый сенсорный центр представлен ядрами таламуса, который является собирателем (коллектором) всех видов сенсорных сигналов за исключением обонятельных.
В процессе проведения происходит пространственное и временнóе преобразование сенсорных сигналов, их переключение, усиление, фильтрация и детектирование. В основе этих функций проводникового отдела лежат такие физиологические процессы как дивергенция возбуждений, конвергенция возбуждений, облегчение, окклюзия, реверберация и процессы торможения в ЦНС.
В корковый (центральный) отдел анализатора поступает наиболее важная – приоритетная информация. Функционально он представлен перцептивными, ассоциативными и гностическими анализаторными зонами коры больших полушарий головного мозга, которые обеспечивают высшие формы анализа и синтеза сенсорных сигналов.
Перцептивные зоны участвуют в формировании простых ощущений. Перцептивная зона соматовисцеральной чувствительности у человека находится, преимущественно, в постцентральной извилине коры больших полушарий. Слуховая область расположена в височной доле коры, а зрительная локализуется в затылочной области. Раздражение этих участков может вызвать соответствующие ощущения прикосновения, звука или света.