Белки мембраны и их функции
Доля белка в общей массе мембраны может колебаться в очень широких пределах – от 18% в миелине до 75% в митохондриальной мембране.
По расположению в мембране белки можно разделить на: интегральные и периферические.
Интегральные белки являются, как правило, гидрофобными и легко встраиваются в липидный бислой.
|
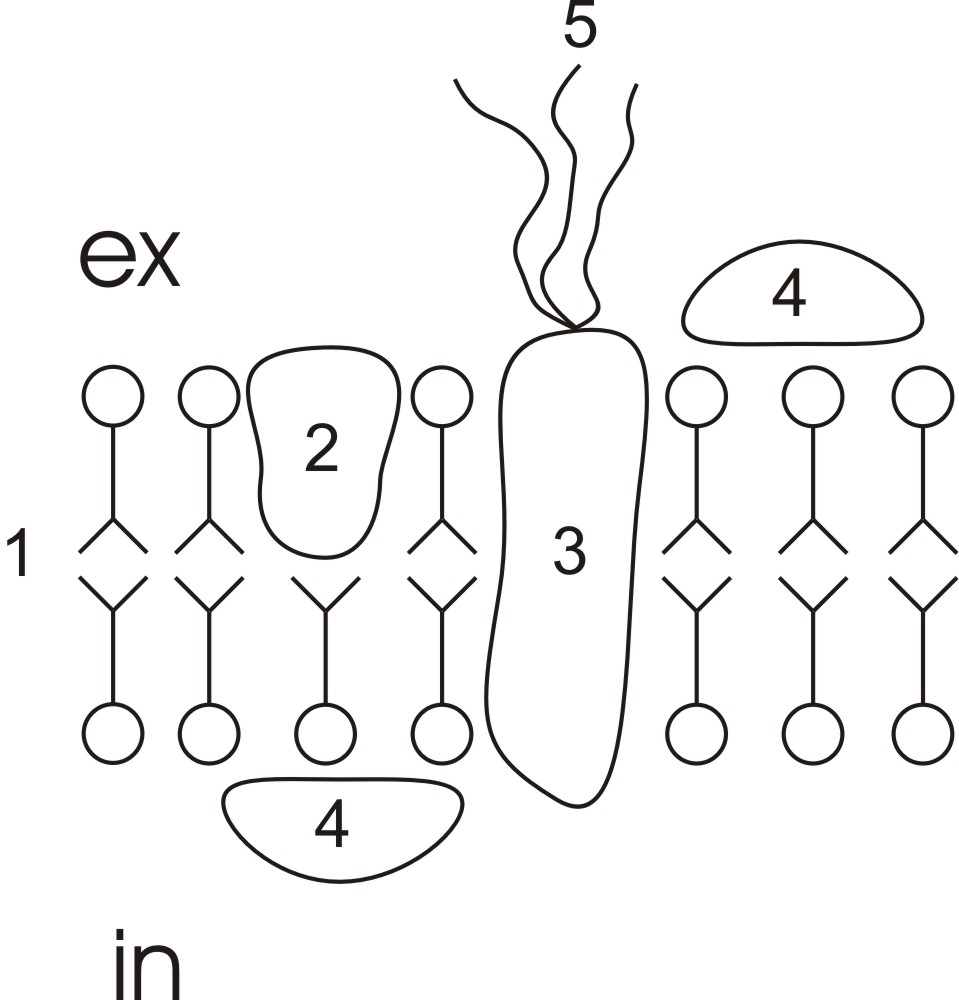
Рис. 9. Классификация белков по месту их расположения в мембране. 1-периферические. 2-интегральные; 3-полуинтегральные.
|

Взаимодействие такого белка с мембраной происходит в несколько стадий. Сначала белок адсорбируется на поверхности бислоя, изменяет свою конформацию, устанавливая гидрофобный контакт с мембраной. Затем происходит внедрение белка в бислой. Глубина внедрения зависит от силы гидрофобного взаимодействия и соотношения гидрофобных и гидрофильных участков на поверхности белковой глобулы. Гидрофильные участки белка взаимодействуют с примембранными слоями по одну или обе стороны мембраны. Фиксация белковой глобулы в мембране происходит благодаря электростатическим и гидрофобным взаимодействиям. Углеводная часть белковых молекул (если она имеется) выступает наружу. Интегральные белки в силу тесной связи с бислоем оказывают на него существенное воздействие: конформационные перестройки белка приводят к изменению состояния липидов, так называемой деформации бислоя.
Периферические белки обладают меньшей глубиной проникновения в липидный бислой, и, соответственно, более слабо взаимодействуют с липидами мембраны, оказывая, на них гораздо меньшее воздействие, чем интегральные.
По характеру взаимодействия с мембраной белки делятся на монотопические, битопические, политопические:
монотопические белки взаимодействуют с поверхностью мембраны (моно – одним из слоев липидов);
битопические пронизывают мембрану насквозь (би – двумя слоями липидов);
политопические пронизывают мембрану несколько раз (поли- многократное взаимодействие с липидами).
Понятно, что первые относятся к периферическим белкам, а вторые и третьи к интегральным.
Белки мембран можно так же классифицировать по выполняемой ими функции. В связи с этим выделяют структурные белки:
белки – ферменты;
белки – рецепторы;
транспортные белки.
Особую группу составляют белки цитоскелета клетки. Строго говоря, эти белки не являются компонентами мембраны, примыкая к ней с цитоплазматической стороны. Белки цитоскелета входят в состав всех его компонентов: миофиламенты содержат молекулы белка актина; в состав микротрубочек входит белок тубулин, промежуточные филаменты также содерждат более полиморфный белковый комплекс. Цитоскелет не только обеспечивают эластичность мембраны, противостоят изменениям объема клетки, но, по-видимому, участвует в и различных внутри- и внеклеточных механизмах регуляции.
Модель биологических мембран
Первоначальные представления о существовании мембран опирались на физиологические исследования. Первые указания на лимитирующие диффузию свойства клеточной поверхности были получены в середине XIX века К.В.Нагели. Он отметил, что клеточная поверхность является барьером для свободной диффузии красителей внутрь клетки из внеклеточной жидкости. Кроме того, Нагели обнаружил, что клетки проявляют осмотические свойства. На основании этих наблюдений К.В.Нагели предположил, что существует некая плазматическая мембрана.
Используя эритроциты в качестве осмометра, Э.Овертон в конце XIX века выявил тесную взаимосвязь между растворимостью вещества в липидах и его способностью проникать в клетку: чем больше эта растворимость, тем меньший осмотический эффект оказывает вещество. Эти данные явились первым свидетельством того, что мембраны содержат большое количество липидов.
Морфологические данные о существовании клеточной мембраны были получены только после разработки методов приготовления ультратонких срезов тканей, фиксированных химическими методами для проведения электронно-микроскопических исследований. Тонкая структура мембран была исследована с помощью метода замораживания – скалывания.
Первыми авторами, которые предложили модель структурной организации мембраны, были Э. Гортер и Ф. Грендель (1925).
Они экстрагировали липиды из теней эритроцитов и приготовляли из них мономолекулярную плёнку на поверхности воды. Амфифильные молекулы липидов распределялись таким образом, что их полярные головки были погружены в воду, а неполярные хвосты торчали наружу. Пленку диспергированных молекул на поверхности воды аккуратно сжимали в латеральном направлении и измеряли силу сжатия. Резкое возрастание этой силы происходило в момент формирования компактного монослоя.
Оказалось, что площадь, занимаемая монослоем, в 2 раза превышала поверхность эритроцитов, взятых для экстракции. Именно это послужило основанием для создания Э. Гортером и Ф. Гренделем концепции липидного бислоя, которая впоследствии легла в основу всех дальнейших представлений о структуре мембран.
Измерение поверхностного натяжения липидного бислоя показало, что оно гораздо больше, чем в мембране эритроцитов, и снижается при добавлении в бислой белков. На основании этих данных в 1931 году Дж. Ф. Даниэлли предложил модель «сэндвича» или унитарную мембранную модель. По его представлениям, белки покрывают обе поверхности липидного бислоя, связываясь с ним электростатическими силами. Из расчётов выходило, что белки находятся на мембране в фибриллярной форме. Этой модели неплохо соответствует миелин – мембрана леммоцитов.
Робертсон несколько модифицировал модель Даниэлли (1964), предположив, что глобулярные белки находятся на внешней стороне мембраны, а фибриллярные белки на внутренней.
Однако с термодинамической точки зрения маловероятно, что белок может успешно конкурировать с водой за полярные головки липидных молекул и что слой белка смог бы экранировать их от водного окружения.
Современные методы исследования окончательно опровергли «бутербродную модель» мембраны.
На сегодняшний день общепризнанной является «мозаичная» модель мембраны, предложенная в 1972 году С. Сингером и Г. Николсоном. Основанием для создания жидкостно-мозаичной модели мембраны послужили данные следующих исследований. Оптические наблюдения показали, что мембранные белки имеют глобулярную структуру. Было установлено, что некоторые белковые молекулы свободно диффундируют в латеральном направлении, т.е. в плоскости мембраны. Исследования с использованием изотопов и др. показали, что белковые молекулы или их части, экспонированные с одной стороны мембраны, отличаются от других, выходящих на другую сторону мембраны.
Согласно этой модели, глобулярные белки интегрированы в липидный бислой; при этом одни из них пронизывают его насквозь, другие лишь частично погружены в бислой. Мембрана является лабильной структурой, все её компоненты имеют возможность осуществлять различные формы подвижности – латеральную диффузию, вращательные движения, «флип – флоп» переходы и другие.
Жидкостно-мозаичная модель, по-видимому, дает наиболее адекватные представления о структурной организации поверхностной мембраны и многих внутриклеточных мембран.
|
Рис. 10. Жидкостно-мозаичная модель биологической мембраны. 1 - липидный бислой; 2 - полуинтегральные белки; 3- интегральные белки; 4- периферические белки; 5 - углеводы.
|