

33. Потенциал действия – изменение проницаемости мембраны для ионов Na и k при генерировании потенциального действия.
Потенциал действия – это общее изменение разности потенциалов на мембране, происходит при возбуждении клетки. Опыты показывают, что возбужденный участок становится электроотрицательным по отношению к покоящемуся, что является показателем перераспределения ионных потоков в возбужденном участке. Для клетки характерен запаздывающий потенциал, когда в течение некоторого времени на мембране существует меньший потенциал, чем потенциал покоя. Схема 1.
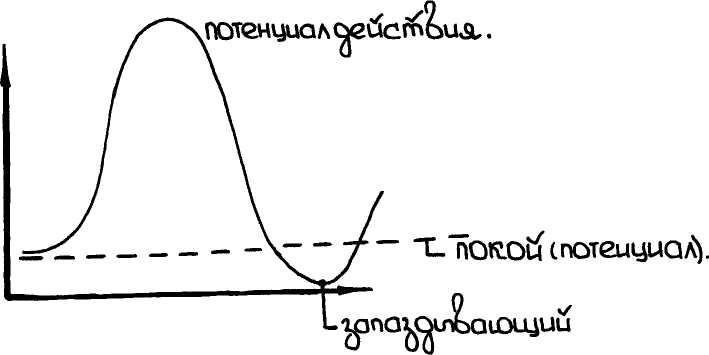
Потенциал действия приводит к кратковременному возрастанию потенциала внутри клетки. Потенциал становится положительным. При возбуждении клетки в начальный период увеличивается проницаемость мембран именно для ионов Na (Хаксли 1963г. на волокнах кальмара) он смог создать опыт с фиксацией определенного значения разности потенциалов на мембране, что позволило ему провести измерения ионных токов и проницаемость (сопротивление) мембран для ионов. Были получены кривые временной зависимости ионных токов через мембрану. Последовательность событий, происходит на клеточной мембране при возбуждении:
1). При возбуждении открываются каналы для ионов Na (проницаемость возрастает в 500 раз) отсюда следует, что отрицательный заряд с внутренней стороны мембраны становится положительным (фаза деполяризации мембраны) – пик мембранного потенциала.
2). Na поступает из внешней среды внутрь, что приводит к тому, что натриевые каналы закрываются (это происходит при генерации импульса) и открываются калиевые. K проходит в соответствие с градиентом концентрации из клетки до тех пор, пока не восстанавливается первоначальный отрицательный заряд на мембране и мембранный потенциал не достигнет своего первоначального значения (фаза реполяризации). За пиком потенциала действия следует небольшой минимум (запаздывающий потенциал).
34. Потенциал зависимые ионные каналы мембраны для k и Na. Структура, особенности функции. Изменение проницаемости мембраны для k и Na в различные фазы потенциального действия.
Входящие и выходящие потоки Na в Na – канале независимы. Это соответствует энергетическому профилю с высоким центральным барьером. Протонирование кислотной группы приводит к блокированию Na – канала. При физиологических значениях pH состоянию канала с депротонированной кислотной группой больше соответственно профиль, в котором центральный энергетический барьер расщепляется на два. В таком канале фиксируют анионный центр, притягивая катионы, облегчая их вход в канал. Однако тем самым одновременно затрудняется выход из основной потенциальной ямы, которая сможет облегчится, когда в боковом участке связывания появляется второй канал. Электростатическое отталкивание способствует выходу первого иона, и второй ион может заполнить центральную яму. Через Na – канал могут проходить различные органические катионы, размеры которого не превышают 0,3 · 0,5 нм. Na – канал открывается для гидроксиламмония и непроницаем для метил аммония (так как вдоль стенок располагаются атомы кислорода и атом водорода OH – группы гидроксиламмоний образует водородную связь, что уменьшает размер молекулы). Энергетический профиль для K – канала: Такой профиль соответствует условиям, когда наиболее медленной стадией транспорта является переход частиц через границу мембраны. Канал все время находится в заполненном двух частичном состоянии. Высокая эффективность ионов – блокаторов с длинной гидрофобной цепью свидетельствует о наличии в устье канала гидрофобных участков. Узкая часть поры (0,26 – 0,3 нм.). непроницаема, как для больших, так и для малых частиц. (небольшой эффект взаимодействия с атомами кислорода, в стенках поры). K – каналы лучше различают ионы, чем Na – каналы.