

[Править]Участвующие в регуляции генов
Основная статья: РНК-интерференция
В живых клетках обнаружено несколько типов РНК, которые могут уменьшать степень выражения гена при комплементарности мРНК или самому гену. Микро-РНК (21-22 нуклеотида в длину) найдены у эукариот и оказывают воздействие через механизм РНК-интерференции. При этом комплекс микро-РНК и ферментов может приводить к метилированию нуклеотидов в ДНК промотора гена, что служит сигналом для уменьшения активности гена. При использовании другого типа регуляции мРНК, комплементарная микро-РНК, деградируется[35]. Однако есть и миРНК, которые увеличивают, а не уменьшают экспрессию генов[36]. Малые интерферирующие РНК (миРНК, 20-25 нуклеотидов) часто образуются в результате расщепления вирусных РНК, но существуют и эндогенные клеточные миРНК[37]. Малые интерферирующие РНК также действуют через РНК-интерференцию по сходным с микро-РНК механизмам[38]. У животных найдены так называемыме РНК, взаимодействующие с Piwi (piRNA, 29-30 нуклеотидов), действующие в половых клетках против транспозиции и играющие роль в образованиигамет[39][40]. Кроме того, piRNA могут эпигенетически наследоваться по материнской линии, передавая потомству своё свойство ингибировать экспрессию транспозонов[41].
Антисмысловые РНК широко распространены у бактерий, многие из них подавляют выражение генов, но некоторые активируют экспрессию[42]. Действуют антисмысловые РНК, присоединяясь к мРНК, что приводит к образованию двуцепочечных молекул РНК, которые деградируются ферментами.[43]. У эукариот обнаружены высокомолекулярные, мРНК-подобные молекулы РНК. Эти молекулы также регулируют выражение генов[44]. В качестве примера можно привести Xist, присоединяющуюся и инактивирующую одну из двух Х-хромосом у самок млекопитающих[45].
Кроме роли отдельных молекул в регуляции генов, регуляторные элементы могут формироваться в 5' и 3' нетранслируемых участках мРНК. Эти элементы могут действовать самостоятельно, предотвращая инициацию трансляции, либо присоединять белки, например, ферритин или малые молекулы, например, биотин[46].
[Править]в процессинге рнк
Основные статьи: Биосинтез белка, Сплайсосома, малые ядерные РНК
Многие РНК принимают участие в модификации других РНК. Интроны вырезаются из пре-мРНК сплайсосомами, которые, кроме белков, содержат несколько малых ядерных РНК (мяРНК)[31]. Кроме того, интроны могут катализировать собственное вырезание[47]. Синтезированая в результате транскрипции РНК также может быть химически модифицирована. У эукариот химические модификации нуклеотидов РНК, например, их метилирование, выполняется малыми ядерными РНК (мяРНК, 60-300 нуклеотидов). Этот тип РНК локализуется вядрышко и тельцах Кахаля[30]. После ассоциации мяРНК с ферментами, мяРНК связываются с РНК-мишенью путём образования пар между основаниями двух молекул, а ферменты модифицируют нуклеотиды РНК-мишени. Рибосомальные и транспортные РНК содержат много подобных модификаций, конкретное положение которых часто сохраняется в процессе эволюции. Также могут быть модифицированы мяРНК и сами мяРНК[48][49]. Гидовые РНК осуществляют процесс редактирования РНК в кинетопласте — особом участке митохондрии протистов-кинетопластид (например, трипаносом).
Рибонуклеи́новая кисло́та (РНК) — одна из трёх основных макромолекул (две другие — ДНК и белки), которые содержатся в клетках всех живых организмов.
Так же, как ДНК (дезоксирибонуклеиновая кислота), РНК состоит из длинной цепи, в которой каждое звено называется нуклеотидом. Каждый нуклеотид состоит из азотистого основания, сахара рибозы и фосфатной группы. Последовательность нуклеотидов позволяет РНК кодироватьгенетическую информацию. Все клеточные организмы используют РНК (мРНК) для программирования синтеза белков.
Клеточные РНК образуются в ходе процесса, называемого транскрипцией, то есть синтеза РНК на матрице ДНК, осуществляемого специальными ферментами — РНК-полимеразами. Затем матричные РНК (мРНК) принимают участие в процессе, называемом трансляцией. Трансляция — это синтез белка на матрице мРНК при участии рибосом. Другие РНК после транскрипции подвергаются химическим модификациям, и после образования вторичной и третичной структур выполняют функции, зависящие от типа РНК.
Для одноцепочечных РНК характерны разнообразные пространственные структуры, в которых часть нуклеотидов одной и той же цепи спарены между собой. Некоторые высокоструктурированные РНК принимают участие в синтезе белка клетки, например, транспортные РНК служат для узнавания кодонов и доставки соответствующих аминокислот к месту синтеза белка, а рибосомные РНК служат структурной и каталитическойосновой рибосом.
Однако функции РНК в современных клетках не ограничиваются их ролью в трансляции. Так, малые ядерные РНК принимают участие всплайсинге эукариотических матричных РНК и других процессах.
Помимо того, что молекулы РНК входят в состав некоторых ферментов (например, теломеразы), у отдельных РНК обнаружена собственнаяферментативная активность: способность вносить разрывы в другие молекулы РНК или, наоборот, «склеивать» два РНК-фрагмента. Такие РНК называются рибозимами.
Геномы ряда вирусов состоят из РНК, то есть у них она играет роль, которую у высших организмов выполняет ДНК. На основании разнообразия функций РНК в клетке была выдвинута гипотеза, согласно которой РНК — первая молекула, которая была способна к самовоспроизведению в добиологических системах.
«11» РЕПЛИКАЦИЯ (от позднелат. replicatio - повторение) (редупликация), самовоспроизведение нуклеиновых к-т (обычно ДНК, у нек-рых вирусов РНК), обеспечивающее точное копирование генетич. информации и передачу ее от поколения к поколению. При репликации ДНК нуклеотидная последовательность копируется (целиком или частично) в виде комплементарной последовательности (см. Комплементарность)дезоксирибонуклеотидов.
В процессе репликации двойная спираль ДНК, состоящая из двух комплементарных полинуклеотидных цепей, раскручивается на отдельные цепи и одновременно начинается синтез новых полинуклеотидных цепей; при этом исходные цепи ДНК играют роль матриц. Новая цепь, синтезирующаяся на каждой из исходных цепей, идентична др. исходной цепи. Когда процесс завершается, образуются две идентичные двойные спирали, каждая из к-рых состоит из одной старой (исходной) и одной новой цепи (рис. 1). Таким образом от одного поколения к другому передается только одна из двух цепей, составляющих исходную молекулу ДНК,-т. наз. полуконсервативный механизм репликации.
Репликация состоит из большого числа последоват. этапов, к-рые включают узнавание точки началу репликации, расплетание исходного дуплекса(спирали), удержание его цепей в изолированном друг от друга состоянии, инициацию синтеза на них новых дочерних цепей, их рост (элонгацию), закручивание цепей в спираль и терминацию (окончание) синтеза. Все эти этапы репликации, протекающие с высокой скоростью и исключит. точностью, обеспечивает комплекс, состоящий более чем из 20 ферментов и белков,-т. наз. ДНК-репликазная система, или реплисома. Функцион. единица репликации-реплик он, представляющий собой сегмент (участок) хромосомы или внехромосомной ДНК, ограниченный точкой начала, в к-рой инициируется репликация, и точкой окончания, в к-рой репликация останавливается. Скорость репликации контролируется на стадии инициации. Однажды начавшись, репликация продолжается до тех пор, пока весь репликон не будет дуплицирован (удвоен). Частотд инициации определяется взаимод. спец. регуляторных белков с точкой начала репликации. Бактериальные хромосомы содержат один репликон: инициации в единств. точке начала репликации ведет к репликации всего генома. В каждом клеточном цикле репликация инициируется только один раз,Плазмиды и вирусы, являющиеся автономными генетич. элементами, представляют собой отдельные репликоны, способные к многократной инициации в клетке-хозяине. Эукариотич. хромосомы (хромосомы всех организмов, за исключением бактерий и синезеленых водорослей) содержат большое число репликонов, каждый из к-рых также однократно инициируется за один клеточный цикл.
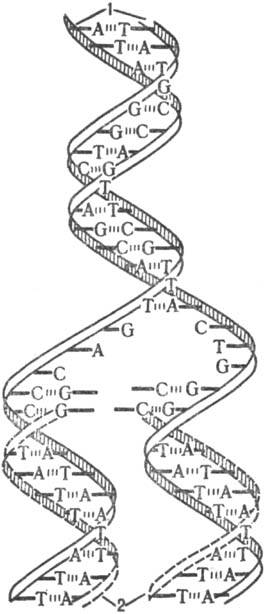
Рис. 1. Схема полуконсервативного механизма репликации: А, Т, G и С-остатки пуриновых и пиримидиновых оснований (соотв. аденина, тимина,гуанина и цитозина); 1 -исходная цепь ДНК; 2-новая цепь ДНК.
Начиная с точки инициации, репликация осуществляется в ограни-ченной зоне, перемещающейся вдоль исходной спирали ДНК. Эта активная зона репликации (т. наз. репликац. вилка) может двигаться в обоих направлениях. При однонаправленной репликации вдоль ДНК движется одна репликац. вилка. При двунаправленной репликации от точки инициации в противоположных направлениях расходятся две репликац. вилки; скорости их движения могут различаться. При репликации ДНК бактерии и млекопитающих скорость роста дочерней цепи составляет соотв. 500 и 50нуклеотидов в 1 с; у растений эта величина не превышает 20 нуклеотидов в 1 с. Движение двух вилок в противоположных направлениях создает петлю, к-рая имеет вид "пузыря" или "глаза". Продолжающаяся репликация расширяет "глаз" до тех пор, пока он не включит в себя весь репликон.
В ходе репликации рост цепи осуществляется благодаря взаимод. дезоксирибонуклеозидтрифосфата с 3'-ОН концевым ну-клеотидом уже построенной части ДНК; при этом отщепляется пирофосфат и образуется фосфодиэфирная связь. Рост полинуклеотидной цепи (рис. 2) идет только с ее З'-конца, т. е. в направлении 5' : 3' (см. Нуклеиновые кисло-ты). Фермент, катализирующий эту р-цию,-ДНК-полиме-раза (см.Полидезоксирибонуклеотид-синтетазы)-не способен начать матричный синтез на одноцепочечной ДНК, если нет хотя бы олигонуклеотидного биспирального участка (т. наз. затравочного олигонуклеотида) комплементарного матрице; затравочным олигонуклеотидом во мн. случаях является не ДНК, а РНК.
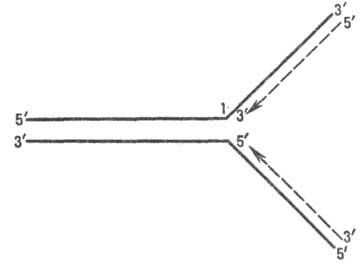
Рис. 2. Направление роста дезоксирибонуклеотидных цепей при репликации; сплошные линии - исходная ДНК, пунктирные - новые цепи ДНК(стрелки показывают направлениеих роста); 1-репликац. вилка.
Энергия, затрачиваемая на образование каждой новой фосфодиэфирной связи в цепи ДНК, обеспечивается расщеплением фосфатной связи между- и -фосфатными группами нуклеозидтрифосфата.
ДНК-полимераза имеет один центр связывания нуклеозидтрифосфата, общий для всех четырех нуклеотидов. Выбор из среды нуклеотида, основаниек-рого комплементарно очередному основанию матрицы, протекает без ошибок, благодаря определяющему влиянию ДНК-матрицы (исходной цепиДНК). При нек-рых мутационных повреждениях структуры ДНК-полимеразы в ряде случаев происходит включение некомплементарных нуклеотидов.
В процессе репликации формальной ДНК на короткое время с вероятностью 10-4-10-5 возникают редкие таутомерные формы всех 4 азотистых оснований нуклеотидов, к-рые образуют неправильные пары. Высокая точность репликации (вероятность ошибок не превышает 10-9) обусловлена наличием механизмов, осуществляющих коррекцию (репарацию).
Репликац. вилка асимметрична. Из двух синтезируемых дочерних цепей ДНК одна строится непрерывно, а другая-с перерывами. Первую наз. ведущей, или лидирующей, цепью, а вторую-отстающей. Синтез второй цепи идет медленнее; хотя в целом эта цепь строится в направлении 3' : 5', каждый из ее фрагментов в отдельности наращивается в направлении 5' : 3' (рис. 3). Благодаря такому прерывистому механизму синтеза, репликация обеих антипараллельных цепей осуществляется с участием одного фермента-ДНК-полимеразы, катализирующего наращивание нуклеотидной цепи только в направлении 5' : 3'.
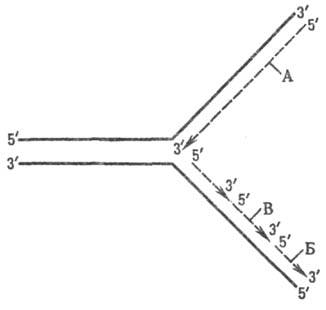
Рис. 3. Схема механизма роста цепей ДНК при репликации: А-ведущая цепь, Б-отстающая цепь, В-фрагмент Оказаки.
В качестве затравок для синтеза фрагментов отстающей цепи служат короткие отрезки РНК, комплементарные матричной цепи ДНК. Эти РНК-затравки (праймеры), состоящие примерно из 10 нуклеотидов, с определенными интервалами синтезируются на матрице отстающей цепи из рибонуклеозидтрифосфатов в направлении 5' : 3' с помощью фермента РНК-праймазы. РНК-праймеры затем наращиваются дезоксинуклеотидами с 3'-конца ДНК-поли-меразой, к-рая продолжает наращивание до тех пор, пока строящаяся цепь не достигает РНК-затравки, присоединенной к 5'-концу предыдущего фрагмента. Образующиеся таким образом фрагменты (т. наз. фрагменты Оказаки) отстающей цепи насчитывают у бактерий 1000-2000 дез-оксирибонуклеотидных остатков; в животных клетках их длина не превышает 200 нуклеотидов.
Чтобы обеспечить образование непрерывной цепи ДНК из многих таких фрагментов, в действие вступает особая система репарации ДНК, удаляющая РНК-затравку и заменяющая ее на ДНК. У бактерий РНК-затравка удаляется нуклеотид за нуклеотидом благодаря 5' : 3'-экзонуклеазнойактивности ДНК-полимеразы. При этом каждый отщепленный рибонуклеотидный мономер замещается соответствующим дезоксирибонуклеотидом (в качестве затравки используется З'-конец синтезированного на старой цепи фрагмента). Завершает весь процесс фермент ДНК-лигаза, катализирующий образование фосфодиэфирной связи между группой З'-ОН нового фрагмента ДНК и 5'-фосфатной группой предыдущего фрагмента. Образование этой связи требует затраты энергии, к-рая поставляется в ходе сопряженного гидролиза пирофосфатной связи кофермента-никотинамид-адениндинуклеотида (в бактериальных клетках) или АТФ (в животных клетках и у бактериофагов).
Раскручивание двойной спирали и пространств. разделение цепей осуществляется при помощи неск. спец. белков. Т. наз. геликазы расплетают короткие участки ДНК, находящиеся непосредственно перед репликац. вилкой. На разделение каждой пары оснований расходуется энергиягидролиза двух молекул АТФ до аденозиндифосфата и фосфата. К каждой из разделившихся цепей присоединяется неск. молекул ДНК-связывающихбелков, к-рые препятствуют образованию комплементарных пар и обратному воссоединению цепей. Благодаря этому нуклеотидные последовательности цепей ДНК оказываются доступными для репликативной системы. Др. специфич. белки помогают праймазе получить доступ кматрице отстающей цепи. В результате праймаза связывается с ДНК и синтезирует РНК-затравки для фрагментов отстающей цепи. Для формирования новых спира-,лей не требуется ни затрат энергии, ни участия к.-л. "закручивающего" фермента.
В случае кольцевого репликона (напр., у плазмиды) описанный процесс наз. -репликацией. Т.к. кольцевые молекулы ДНК закручены сами на себя (суперспирализо-ваны), при раскручивании двойной спирали в процессе репликации они должны непрерывно вращаться вокруг собств. оси. При этом возникает торсионное напряжение, к-рое устраняется путем разрыва одной из цепей. Затем оба конца сразу же вновь соединяются друг с другом. Эту ф-цию выполняет фермент ДНК-топоизомераза. Репликация в этом случае обычно происходит в двух направлениях, т.е. существуют две репликац. вилки (рис. 4). После завершения репликации появляются две двухцепочечные молекулы, к-рые сначала связаны друг с другом как звенья одной цепи. При их разделении одно из двух колец временно разрывается.

Рис. 4. Один из механизмов репликации плазмиды (начало репликации обозначено точками); направления движения репликац. вилки показаны стрелками, образующиеся новые цепи ДНК-пунктиром.
Альтернативный вариант репликации кольцевого репликона предполагает разрыв в одной из цепей двухспиральной молекулы ДНК. Образовавшийся при этом свободный 3'-конец кова-лентно наращивается, оставаясь связанным с матрицей (второй, неразорванной цепью), а 5'-конец постепенно вытесняется новой полинуклеотидной цепью (рис. 5). Таким образом одна цепь разматывается и непрерывно удлиняется, а репликац. вилка скользит вокруг кольцевой матричной цепи (механизм "катящегося кольца"). По мере роста новой цепи вытесненная цепь с освободившимся 5'-концом становится линейной матрицей для синтеза новой комплементарной цепи. Этот синтез на линейной матрице продолжается до тех пор, пока не образуется дочерняя цепь ДНК, комплементарная одному обороту кольцевой матрицы, т. е. целому репликону. Таким путем с кольцевойматрицы может сходить большое число комплементарных копий. Такой механизм обнаружен у нек-рых вирусов, а также в ряде клеток эукариот.
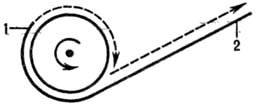
Рис. 5. Схема репликации по механизму катящегося кольца (новая молекула ДНК показана пунктиром): 1-3'-конец ДНК; 2-5'-конец ДНК.
Еще одна схема репликации предполагает формирование структуры, названной D-петлей. Согласно этому механизму, сначала реплицируется только одна из цепей кольцевого репликона, тогда как вторая цепь, оставаясь интактной, вытесняется, образуя петлю. Репликация второй цепи начинается с др. стартовой точки и только после того, как реплицировалась часть первой цепи. Такой механизм репликации обнаружен, напр., у митохондриальных ДНК.
Репликация РНК (синтез РНК на РНК-матрице) изучена меньше. Она осуществляется только у нек-рых вирусов (напр., у вирусов полиомиелита и бешенства). Фермент, катализирующий этот процесс,-РНК-зависимая РНК-полимераза (его называют также РНК-репликазой или РНК-синтетазой). Известно неск. типов репликации, РНК: 1) вирусы, содержащие матричные РНК, или мРНК [т. наз. (+)РНК], в результате репликации образуют комплементарную ей цепь [(-)РНК], не являющуюся мРНК, к-рая используется как матрица для синтеза (+)РНК; 2) вирусы, содержащие (—)РНК, в результате репликации синтезируют (+)РНК; 3) вирусы, содержащие двухцепочечную РНК [(+)PHK и (—)РНК], в результате асимметрической репликации синтезируют (+)РНК.
Гипотеза о механизме репликации сформулирована в 1953 Дж. Уотсоном и Ф. Криком, к-рые предположили, что две комплементарные цепи ДНКпосле их разделения могут выполнять ф-ции матриц для образования на них новых цепей ДНК. В 1958 М. Мезельсон и Ф. Сталь экспериментально подтвердили такой механизм репликации.
«12» ТРАНСКРИПЦИЯ (от лат. transcriptio, букв-переписывание), биосинтез РНК на матрице ДНК; первая стадия реализации генетич. информации, в ходе к-рой нуклеотидная последовательность ДНК считывается в виде нуклеотидной последовательности РНК (см. Генетический код). В основе этого процесса лежит принцип комплементарного спаривания пуриновых и пиримидиновых оснований (см. Компле-ментарность). Транскрипция осуществляется с участием фермента РНК-полимеразы, использующей в качестве субстратов рибонук-леозидтрифосфаты. Кроме того, в транскрипции участвует большое число вспомогат. белков, регулирующих работу РНК-полимеразы.
Транскрипция происходит на участках ДНК, наз. единицами транскрипции или трапскриптонами. В начале и конце транскрилтона расположены специфич. нуклеотидные последовательности -соотв. промотор и терминатор. Существование множества транскриптонов обеспечивает возможность независимого считывания разных генов, их индивидуального включения и выключения. У животных, растений и др. эукариот в составтранскриптона, как правило, входит один ген. Транс-криптоны бактерий обычно наз. оперонами; мн. из них содержат по неск. генов, обычно функционально связанных (напр., кодирующих неск. ферментов, участвующих в синтезе той или иной аминокислоты).
Процесс синтеза РНК можно разделить на четыре основные стадии: 1) связывание РНК-полимеразы с промотором, 2) начало синтеза цепи РНК(инициация), 3) рост цепи РНК (элонгация), 4) завершение синтеза цепи РНК (терминация).
Связывание РНК-полимеразы с промотором включает по крайней мере два этапа. На первом РНК-полимераза образует с промотором закрытый комплекс, в к-ром ДНК сохраняет двухспиральную структуру, а РНК-полимераза еще не способна начать синтез РНК. На втором закрытый комплекс превращается в открытый, в к-ром РНК-полимераза расплетает примерно один виток двойной спирали ДНК в районе стартовой точки-нуклеотида, с к-рого начинается комплементарное копирование матрицы.
При наличии субстратов РНК-полимераза в открытом комплексе осуществляет инициацию. Первый нуклеотид (обычно это аденозин- или гуанозинтрифосфат) входит в состав цепи целиком, а последующие присоединяются к группе 3'-ОН предыдущего нуклеотида с образованием фос-фодиэфирной связи и освобождением пирофосфата (см. Нуклеиновые кислоты). На стадии инициации образующаяся РНК связана с матрицей иферментом непрочно и может отделиться от комплекса. В этом случае РНК-полимераза, не покидая промотора, снова инициирует РНК (такой синтез коротких рибонуклеотидов наз. абортивным). Стадия инициации завершается, когда цепь РНК достигает критич. длины (от 3 до 9 нуклеотидовна разных промоторах); при этом от РНК-полимеразы отделяется -субъединица.
Считают, что в процессе элонгации примерно 13 нуклеотидов РНК образуют гибридную спираль с матричной нитью расплетенной ДНК (всего на этой стадии в ДНК расплетено примерно 18 нуклеотидов). По мере движения РНК-полимеразы по матрице впереди нее происходит расплетание, а позади восстановление двойной спирали ДНК. Одновременно происходит вытеснение очередного звена растущей цепи РНК из комплекса сматрицей.
Цепь РНК растет в направлении 5' : 3' по мере продвижения РНК-полимеразы по цепи ДНК в направлении от 3'-конца к 5'-концу. Средняя скорость роста цепи РНК у бактерии Escherichia coli (E. coli) составляет 40-45 рибонуклеотидов в секунду. В процессе удлинения цепи РНК фермент движется по ДНК с непостоянной скоростью. В нек-рых участках матрицы происходят длительные задержки в его продвижении, т. наз. паузы (нек-рые стадии транскрипции показаны на рис.).
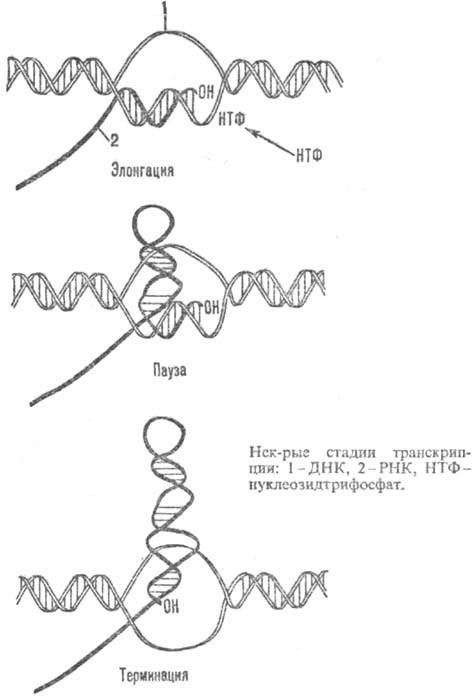
На стадии злонгации в состав транскрибирующего комплекса входит ряд дополнит. белков, от к-рых зависит протекание завершающей стадии транскрипции -терминации. Один из таких белков, кодируемых геном nusA E. coli, занимает в РНК-полимеразе место -субъединицы. Др. бактериальный фактор терминации взаимод. с РНК.
Терминация транскрипции, как правило, происходит в строго определенных участках матрицы - терминаторах, в к-рых от матрицы отделяются РНКи РНК-полимераза; последняя, объединившись со свободной -субъединицей, может вступить в следующий цикл транскрипции. В терминаторах, для узнавания к-рых РНК-полимеразе не требуется фактора р, нуклеотидная последовательность характеризуется двумя особенностями: по ходу транскрипции перед точкой терминации расположен участок, богатый парами dG-dC (дезоксигуанозин-дезоксицити-дин), а затем участок, состоящий из 4-8 расположенных подряд остатков дезоксиадениловой к-ты. Предполагают, что после прохождения РНК-полимеразой участка, богатого dG-dC, в РНК возникает шпилька, к-рая препятствует продвижению фермента и разрушает часть спирали РНК-ДНК транскрибирующего комплекса. Оставшаяся часть гибридной спирали, включающая концевую полиуридиловую последовательность РНК, легко плавится (разрушается) ввиду крайней нестабильности комплементарной пары уридин-дезоксиаденозин, что и приводит к освобождению РНК.
Мн. терминаторы узнаются РНК-полимеразой только с помощью белковых факторов терминации. Из них наиб. изучен фактор E. coli-олигомерныйбелок с мол. м. 46 тыс. Фактор присоединяется к определенным участкам синтезируемой РНК (не имеющим протяженных двухспи-ральных структур) до того, как РНК-полимераза достигает терминатора. Предполагается, что фактор передвигается вдоль РНК вслед за РНК-полимеразой, используя для этого энергию гидролиза нуклеозидтрифосфатов, и способствует диссоциации гибрида РНК с матричной нитью ДНК.
Скорость транскрипции разл. генов может отличаться в тысячи раз; в столь же больших пределах может изменяться скорость транскрипции одного и того же гена в разных тканях многоклеточного организма или в одной клетке в зависимости от изменяющихся внеш. условий или внутр. программы. На стадии инициации регуляция транскрипции осуществляется благодаря наличию особых белков-регуляторов (см. Регуляторные белки), способных присоединяться к определенным участкам ДНК и тем самым препятствовать или помогать РНК-полимеразе инициировать синтез РНК напромоторе.
У прокариот регуляция транскрипции часто осуществляется на стадии терминации в особых терминаторах (называемых аттенюаторами), расположенных в начале или внутри оперонов.
Существует также обратная транскрипция-синтез ДНК на матрице РНК. Такой синтез осуществляется у ретровирусов (семейство РНК-содержащихвирусов) с участием фермента ревер-тазы (обратная транскриптаза). В ходе обратной транскрипции образуется вначале гибрид РНК-ДНК, к-рый реплицирует под действием ДНК-зависимой ДНК-полимеразы (см. Полиде-зоксирибонуклеотид-синтетазы) с образованием двухцепо-чечной спиралиДНК. Последняя также подвержена репликации и способна включаться в геном инфицированной клетки и служить там матрицей для вирусной РНК. Транскрипция обр., поток генетич. информации у ретровирусов направлен от РНК к ДНК и затем обратно к РНК.
«13» Созревание РНК
Большинство меток организма содержит полный набор генов, но обычно из этого набора используется крайне незначительный объем информации. Постоянно транскрибируются только те гены, которые кодируют структурные белки и ферменты промежуточного метаболизма. Кроме этих постоянно необходимых генов имеется много других генов, активных только в определенных типах клеток, при определенных метаболических условиях или во время дифференцировки.
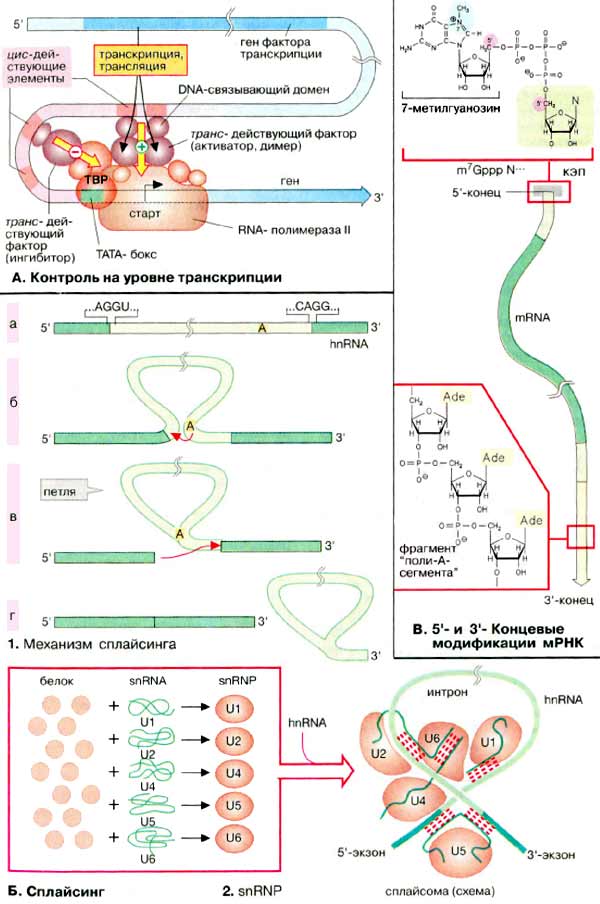
А. Контроль на уровне транскрипции
Порядок транскрибирования генов определяется регуляторной системой, которая носит название системы регуляции транскрипции. Контроль транскрипции осуществляется структурами двух типов. Большинство генов содержат в своем промоторном участке (см. с. 240) несколько коротких сегментов ДНК (DNA) (регуляторные элементы, цис-действующие элементы), с которыми могут связываться факторы транскрипции. Регуляторные элементы, стимулирующие транскрипцию связанных с ними генов, называются энхансерами (усилителями, от англ. enhancer). Белки, подавляющие транскрипцию. — сайленсерами (успокоителями, от англ. silencer). Факторы транскрипции — это белки, т. е. продукты других, независимых генов. Поэтому их называют опосредованно действующими факторами. Для процесса транскрипции генов требуются не только РНК-полимераза, но и другие белки, называемые основными факторами транскрипции. Установлено, что у эукариот таким фактором является ТАТА-связывающий белок (ТСБ, англ. ТАТА-Вох Binding Protein, TBP), который взаимодействует с основным регуляторным элементом. ТАТА-боксом, присутствующим в большинстве генов (см. с. 240). С этим комплексом затем связываются другие основные факторы транскрипции и РНК-полимеразы. Дополнительные факторы могут влиять на инициацию транскрипции, связываясь с другими регуляторными элементами. Отсюда они взаимодействуют с основным транскрипционным комплексом, либо активируя, либо ингибируя его. Такие факторы активируют, например, комплексы стероидных гормонов с рецепторами (см. с. 366). По завершении транскрипции из гяРНК вырезаются интроны (см. с. 240), содержащие некодирующие последовательности.
Б. Сплайсинг
Сплайсинг РНК катализируется комплексами белков с РНК, известными как «малые ядерные рибонуклеопротеидные частицы» (мяРНП, англ. small nuclear ribonucleic particles, snRNP). Интроны, входящие в гяРНК (hnRNA), имеют специфические последовательности на 3'- и 5'-концах (а). На первой стадии сплайсинга ОН-группа аденозилового остатка, расположенного в интроне, атакует (при участии мяРНП) и расщепляет фосфодиэфирную связь на 5'-конце интрона (б). Одновременно в интроне образуется новая связь, которая придает ему форму петли (в). На второй стадии терминальная ОН-группа 5'-концевого интрона атакует связь в 3'-конце интрона. В результате оба экзона соединяются, а интрон освобождается (г).
В этой реакции принимают участие пять различных мяРНП (U1, U2, U4, U5 и U6). В каждой из реакций задействованы несколько белковых молекули одна молекула мяРНК (snRNA) (см. с. 88). Во время сплайсинга комплексы из гяРНК и мяРНП образуют сплайсому. Полагают, что мяРНК в сплайсоме образуют канонические пары друг с другом и с гяРНК и таким образом фиксируют и ориентируют их реакционные группы. Собственнокатализ обусловлен РНК-составляющей сплайсомы Такие каталитические РНК носят название рибозимов.
В. 5'- и 3'-Концевые модификации мРНК
У эукариот после завершения собственно транскрипции 5'-конец растущей молекулы РНК блокируется структурой, которая называется кэп (от англ. cap). В случае мРНК кэп состоит из 7'-метил-ГТФ и защищает РНК от гидролиза 5'-экзонуклеазами. В конце транскрипции к 3'-концу присоединяетсяполиадениловая последовательность, которая может включать до 200 звеньев АМФ (AMP). Только после этого созревшая мРНК (mRNA) покидает ядро.
«14» Рибосома — важнейший немембранный органоид живой клетки сферической или слегка эллипсоидной формы, диаметром от 15—20нанометров (прокариоты) до 25—30 нанометров (эукариоты), состоящий из большой и малой субъединиц. Рибосомы служат для биосинтеза белка из аминокислот по заданной матрице на основе генетической информации, предоставляемой матричной РНК, или мРНК. Этот процесс называется трансляцией.
В эукариотических клетках рибосомы располагаются на мембранах эндоплазматической сети, хотя могут быть локализованы и в неприкрепленной форме в цитоплазме. Нередко с одной молекулой мРНК ассоциировано несколько рибосом, такая структура называетсяполирибосомой (полисомой). Синтез рибосом у эукариот происходит в специальной внутриядерной структуре — ядрышке.
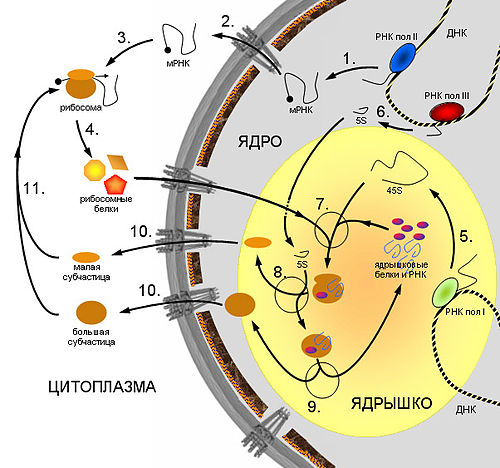
Схема синтеза рибосом в клетках эукариот. 1. Синтез мРНК рибосомных белков РНК полимеразой II. 2. Экспорт мРНК из ядра. 3. Узнавание мРНК рибосомой и 4. синтез рибосомных белков. 5. Синтез предшественника рРНК (45S — предшественник) РНК полимеразой I. 6. Синтез 5S pРНК РНК полимеразой III. 7. Сборка большой рибонуклеопротеидной частицы, включающей 45S-предшественник, импортированные из цитоплазмы рибосомные белки, а также специальные ядрышковые белки и РНК, принимающие участие в созревании рибосомных субчастиц. 8. Присоединение 5S рРНК, нарезание предшественника и отделение малой рибосомной субчастицы. 9. Дозревание большой субчастицы, высвобождение ядрышковых белков и РНК. 10. Выход рибосомных субчастиц из ядра. 11. Вовлечение их в трансляцию.
Рибосомы представляют собой нуклеопротеид, в составе которого отношение РНК/белок составляет 1:1 у высших животных и 60-65:35-40 у бактерий. Рибосомная РНК составляет около 70 % всей РНК клетки. Рибосомы эукариот включают четыре молекулы рРНК, из них 18S, 5.8S и 28S рРНК синтезируются в ядрышке РНК полимеразой I в виде единого предшественника (45S), который затем подвергается модификациям и нарезанию. 5S рРНК синтезируется РНК полимеразой III в другой части генома и не нуждаются в дополнительных модификациях. Почти вся рРНК находится в виде магниевой соли, что необходимо для поддержания структуры; при удалении ионов магния рибосома подвергается диссоциации на субъединицы.
Константа седиментации (скорость оседания в ультрацентрифуге) рибосом эукариотических клеток равняется 80S (большая и малая субъединицы 60S и 40S, соответственно), бактериальных клеток (а также митохондрий и пластид) — 70S (большая и малая субъединицы 50S и 30S, соответственно).