

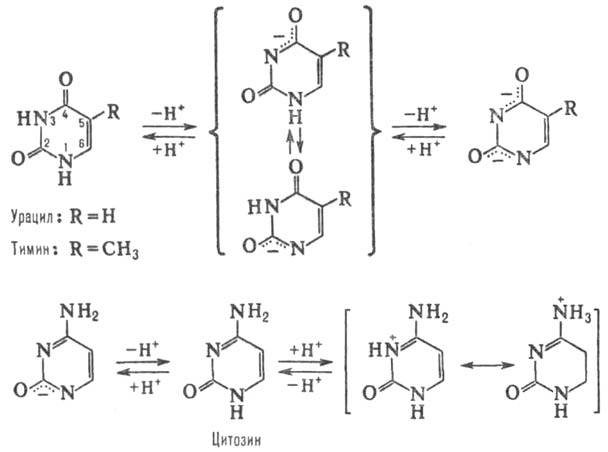
ПИРИМИДИНОВЫЕ ОСНОВАНИЯ, производные пиримидина, входящие в состав нуклеиновых к-т, нуклео-тидов, коферментов и др. Канонич. пиримидиновые основания -цитозин (4-амино-2-пиримидон, сокращенно С), тимин (3-метил-пиримидин-2,4-дион, T) и урацил (пиримидин-2,4-дион, U); разл. формы молекул. Пиримидиновые основания (они существуют при разных значениях рН) показаны на схеме.
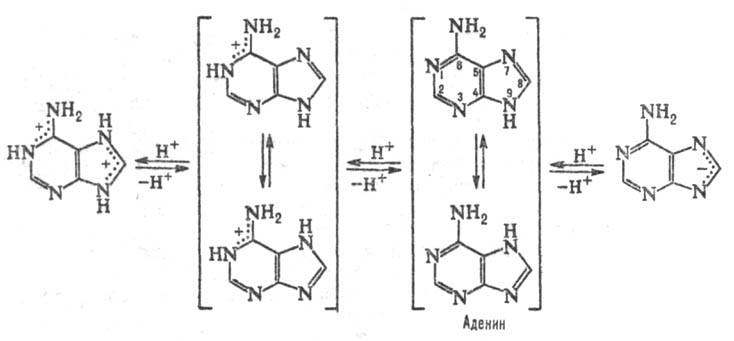
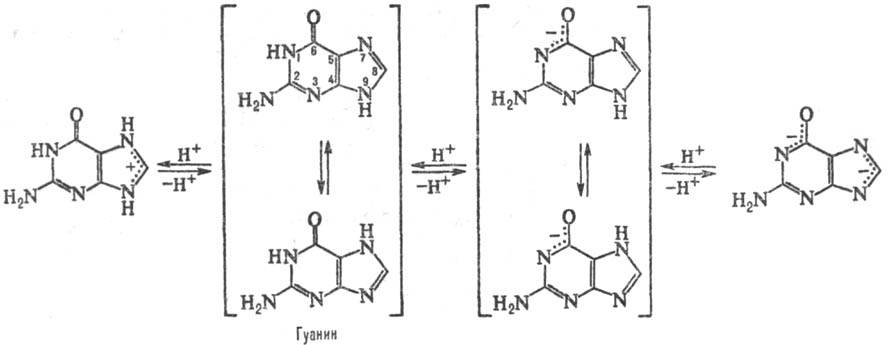
ПУРИНОВЫЕ ОСНОВАНИЯ, прир. производные пурина. Входят в качестве агликонов (неуглеводного компонента) в нуклеиновые к-ты, нуклеозиды,нуклеотиды; фрагменты коферментов, витаминов и др. Канонические пуриновые основания нуклеиновых к-т-аденин (6-аминопурин, сокращенно А) игуанин (2-амино-6-пуринон, G). Разл. формы молекул пуриновых оснований, к-рые существуют при разных значениях рН, и таутомерные формы показаны на схеме:
Строение нуклеотидов
Каждый нуклеотид содержит 3 химически различных компонента: гетероциклическое азотистое основание, моносахарид (пентозу) и остаток фосфорной кислоты. В зависимости от числа имеющихся в молекуле остатков фосфорной кислоты различают нуклеозидмонофосфаты (НМФ), нуклеозиддифосфаты (НДФ), нуклео-зидтрифосфаты (НТФ) (рис. 4-1).
В состав нуклеиновых кислот входят азотистые основания двух типов: пуриновые - аденин (А),гуанин (G) и пиримидиновые - цитозин (С), тимин (Т) и урацил (U). Нумерация атомов в основаниях записывается внутри цикла (рис. 4-2). Номенклатура нуклеотидов приведена в табл. 4-1.
Пентозы в нуклеотидах представлены либо рибозой (в составе РНК), либо дезоксирибозой (в составе ДНК). Чтобы отличить номера атомов в пентозах от нумерации атомов в основаниях, запись производят с внешней стороны цикла и к цифре добавляют штрих (') - 1', 2', 3', 4' и 5' (рис. 4-3).
Пентозу соединяет с основанием N-гликозидная связь, образованная С1-атомом пентозы (рибозы или дезоксирибозы) и N1 -атомом пиримидина или N9-aтомом пурина (рис. 4-4).
Нуклеотиды,
в которых пентоза представлена рибозой,
называют рибонуклеотидами, а нуклеиновые
кислоты, построенные из рибонуклеотидов,
- рибонуклеиновыми кислотами, или РНК.
Нуклеиновые кислоты, в мономеры которых
входит дезоксирибоза, называют
дезоксири-бонуклеиновыми кислотами,
или ДНК. Нуклеиновые кислоты по своему
строению относят кклассу
линейных полимеров. Остов нуклеиновой
кислоты имеет одинаковое строение по
всей длине молекулы и состоит из
чередующихся групп - пентоза-фосфат-пентоза-
(рис. 4-5). Вариабельными группами в
полинуклеотидных цепях служат азотистые
основания - пурины и пиримидины. В
молекулы РНК входят аденин (А), урацил
(U), гуанин (G) и цитозин (С), в ДНК - аденин
(А), тимин (Т), гуанин (G) и цитозин (С).
Уникальность структуры и функциональная
индивидуальность молекул ДНК и РНК
определяются их первичной структурой
-последовательностью азотистых оснований
в полинуклеотидной цепи.
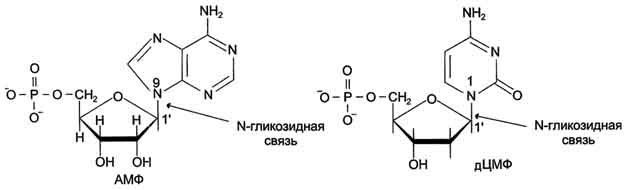
Рис. 4-4. Пуриновый и пиримидиновый нуклеотиды.
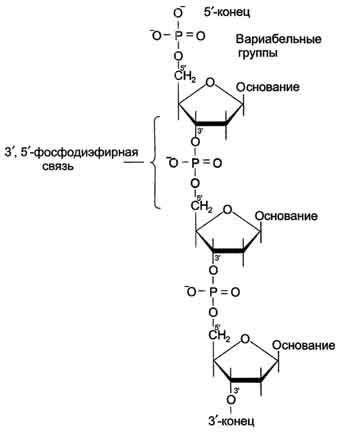
Рис. 4-5. Фрагмент цепи ДНК.
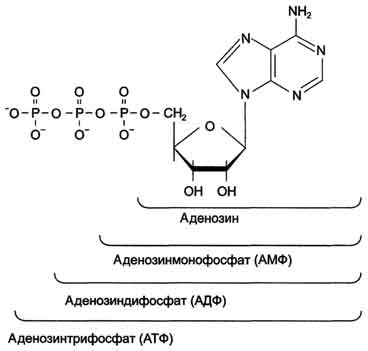
Рис. 4-1. Нуклеозидмоно-, ди- и трифосфаты аденозина. Нуклеотиды - фосфорные эфиры нуклеозидов. Остаток фосфорной кислоты присоединён к 5'-углеродному атому пентозы (5'-фосфоэфирная связь).
141
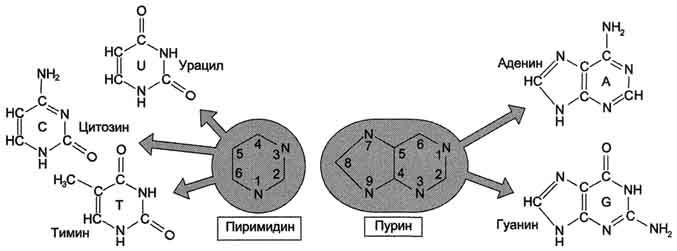
Рис. 4-2. Пуриновые и пиримидиновые основания.
Таблица 4-1. Номенклатура нуклеотидов
Азотистое основание |
Нуклеозид |
Нуклеотид |
Трёхбуквенное обозначение |
Однобуквенный код |
Аденин |
Аденозин |
Аденозинмонофосфат |
АМФ |
А |
Гуанин |
Гуанозин |
Гуанозинмонофосфат |
ГМФ |
G |
Цитозин |
Цитидин |
Цитидинмонофосфат |
ЦМФ |
С |
Урацил |
Уридин |
Уридинмонофосфат |
УМФ |
U |
Тимин |
Тимидин |
Тимидинмонофосфат |
ТМФ |
Т |
«2»
|
Сравнительная характеристика ДНК и РНК
|
||||||||||||||||||
«3» КИСЛОТЫ, ИХ СТРУКТУРА И СВОЙСТВА Нуклеозиды и нуклеотиды являются продуктами гидролиза нуклеиновых кислот, но они присутствуют в живых организмах также в несвязанном состоянии, выполняя исключительно важную роль в обмене веществ.
Нуклеозиды
-
это природные гликозиды гетероциклических
азотистых оснований (пиримидиновых
и пуриновых),
которые связаны с пентозами через
атомазота.
В зависимости от природы углеводного
остатка (пентозы)
различают рибонуклеозиды и
дезоксирибонуклеозиды.
 Пиримидиновые
основания:
Пиримидиновые
основания:
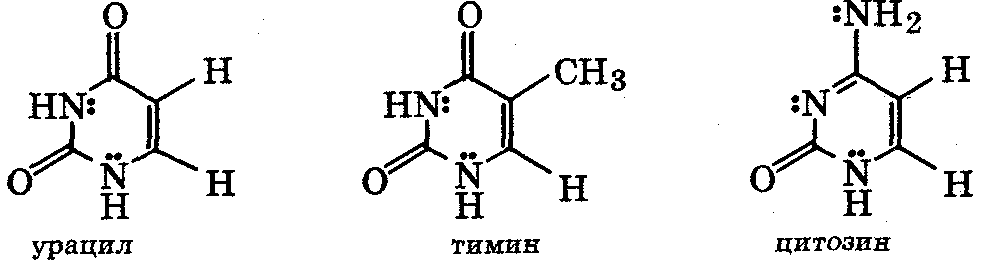 Пуриновые
основания:
Пуриновые
основания:
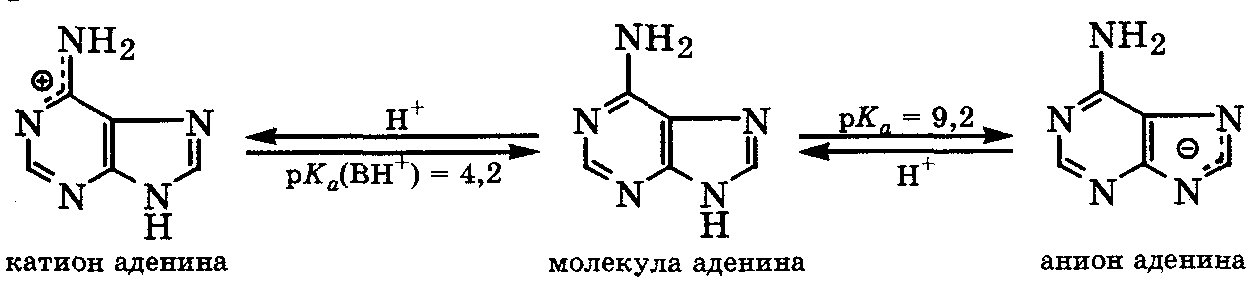
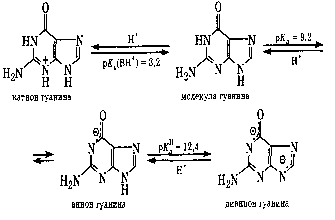 Названия
нуклеозидов
производятся
от тривиального названия соответствующего
гетероциклического азотистого основания
с
суффиксами -идин у пиримидиновых и
-озин у пуриновых нуклеозидов. Исключение их
этого правила сделано для
нук-
леозидов
тимина.
Названия
нуклеозидов
производятся
от тривиального названия соответствующего
гетероциклического азотистого основания
с
суффиксами -идин у пиримидиновых и
-озин у пуриновых нуклеозидов. Исключение их
этого правила сделано для
нук-
леозидов
тимина.
 Нуклеозиды
сокращенно чаще обозначают однобуквенным
индексом, но существует также система
трехбуквенного индекса.
Нуклеозиды,
являясь N-гликозидами,
устойчивы к гидролизу в слабощелочной
среде, но расщепляются в кислой среде.
Пуринонуклеозиды гидролизуются легко,
пирими-диновые - труднее.
Нуклеозиды
сокращенно чаще обозначают однобуквенным
индексом, но существует также система
трехбуквенного индекса.
Нуклеозиды,
являясь N-гликозидами,
устойчивы к гидролизу в слабощелочной
среде, но расщепляются в кислой среде.
Пуринонуклеозиды гидролизуются легко,
пирими-диновые - труднее.
Нуклеотиды
являются сложными
эфирами нуклеозидов и фосфорной кислоты,
которая обычно этерифицирует гидроксогруппы
при С-5' пентозы. В связи с наличием в
молекуле остатка фосфорной кислоты
нуклеотиды проявляют свойства двухосновной
кислоты с pKa1 =
0,9 - 1,5 и рKaП =
6 - 6,5.
 Нуклеотиды называют
или как соответствующие кислоты
(монозамещенные производные фосфорной
кислоты), или как соли (монофосфаты) с
указанием в обоих случаях положения
фосфатного остатка:
Нуклеотиды называют
или как соответствующие кислоты
(монозамещенные производные фосфорной
кислоты), или как соли (монофосфаты) с
указанием в обоих случаях положения
фосфатного остатка:
 Большое
значение в живых системах играют
нуклеотиды, содержащие в своем составе
ди- и трифосфатные группировки. Важнейшими
среди этих производных являются аденозиндифосфат
(АДФ) и аденозинтрифосфат (АТФ), которые
способны к взаимо-превращениям путем
наращивания или отщепления фосфатных
групп:
Большое
значение в живых системах играют
нуклеотиды, содержащие в своем составе
ди- и трифосфатные группировки. Важнейшими
среди этих производных являются аденозиндифосфат
(АДФ) и аденозинтрифосфат (АТФ), которые
способны к взаимо-превращениям путем
наращивания или отщепления фосфатных
групп:
 В
этих соединениях фосфатные группы в
физиологических условиях почти полностью
ионизованы, поэтому их часто записывают
в виде анионов АТФ4-,
АДФ3-.
Главная особенность этих нуклеотидов
состоит в том, что их полифосфатные
группы содержат одну или две ангидридные
группы
—Р—О—Р—
II II
О
О
.
При гидролизе такой группы разрывается
связь, называемая макроэргической,
и выделяется около 33 кДж/моль. Именно с
этим связана роль АТФ в клетке как
поставщика химической энергии для
биохимических и физиологических
процессов.
При
участии АТФ в организме также
осуществляется реакция
фосфорилирования гидроксилсодержащих
соединений с образованием сложных
эфиров фосфорной кислоты:
В
этих соединениях фосфатные группы в
физиологических условиях почти полностью
ионизованы, поэтому их часто записывают
в виде анионов АТФ4-,
АДФ3-.
Главная особенность этих нуклеотидов
состоит в том, что их полифосфатные
группы содержат одну или две ангидридные
группы
—Р—О—Р—
II II
О
О
.
При гидролизе такой группы разрывается
связь, называемая макроэргической,
и выделяется около 33 кДж/моль. Именно с
этим связана роль АТФ в клетке как
поставщика химической энергии для
биохимических и физиологических
процессов.
При
участии АТФ в организме также
осуществляется реакция
фосфорилирования гидроксилсодержащих
соединений с образованием сложных
эфиров фосфорной кислоты:
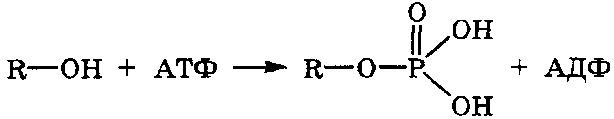 При
фосфорилировании карбоксилсодержащих
соединений образуются ацилфосфаты,
которые содержат ангидридную
группировку:
При
фосфорилировании карбоксилсодержащих
соединений образуются ацилфосфаты,
которые содержат ангидридную
группировку:
![]() –
–![]() С—О—Р
׀׀
׀׀
О
О
С—О—Р
׀׀
׀׀
О
О
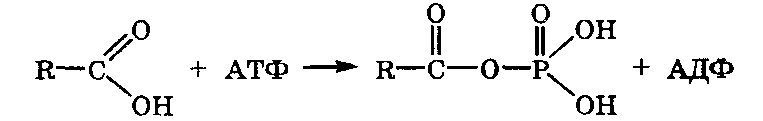 Фосфорилированные
производные выступают активными
метаболитами во многих биохимических
процессах.
. Важнейшая
биологическая роль нуклеотидов
заключается в том, что из них построены
нуклеиновыекислоты (полинуклеотиды).
Фосфорилированные
производные выступают активными
метаболитами во многих биохимических
процессах.
. Важнейшая
биологическая роль нуклеотидов
заключается в том, что из них построены
нуклеиновыекислоты (полинуклеотиды).
«4» Аденозинтрифосфа́т (сокр. АТФ, англ. АТР) — нуклеотид, играет исключительно важную роль в обмене энергии и веществ в организмах; в первую очередь соединение известно как универсальный источник энергии для всех биохимических процессов, протекающих в живых системах. АТФ был открыт в 1929 году Карлом Ломанном[1], а в 1941 году Фриц Липман показал, что АТФ является основным переносчиком энергии в клетке[2]. Систематическое наименование АТФ:
9-β-D-рибофуранозиладенин-5'-трифосфат, или
9-β-D-рибофуранозил-6-амино-пурин-5'-трифосфат.
Химически АТФ представляет собой трифосфорный эфир аденозина, который является производным аденина и рибозы.
Пуриновое азотистое основание — аденин — соединяется β-N-гликозидной связью с 1'-углеродом рибозы. К 5'-углероду рибозы последовательно присоединяются три молекулыфосфорной кислоты, обозначаемые соответственно буквами: α, β и γ.
АТФ относится к так называемым макроэргическим соединениям, то есть к химическим соединениям, содержащим связи, при гидролизе которых происходит освобождение значительного количества энергии. Гидролиз макроэргических связей молекулы АТФ, сопровождаемый отщеплением 1 или 2 остатков фосфорной кислоты, приводит к выделению, по различным данным, от 40 до 60 кДж/моль.
АТФ + H2O → АДФ + H3PO4 + энергия
АТФ + H2O → АМФ + H4P2O7 + энергия
Высвобожденная энергия используется в разнообразных процессах, протекающих с затратой энергии.
[Править]Роль в организме
Главная роль АТФ в организме связана с обеспечением энергией многочисленных биохимических реакций. Являясь носителем двух высокоэнергетических связей, АТФ служит непосредственным источником энергии для множества энергозатратных биохимических и физиологических процессов. Всё это реакции синтеза сложных веществ в организме: осуществление активного переноса молекул черезбиологические мембраны, в том числе и для создания трансмембранного электрического потенциала; осуществления мышечного сокращения.
Помимо энергетической АТФ выполняет в организме ещё ряд других не менее важных функций:
Вместе с другими нуклеозидтрифосфатами АТФ является исходным продуктом при синтезе нуклеиновых кислот.
Кроме того, АТФ отводится важное место в регуляции множества биохимических процессов. Являясь аллостерическим эффектором ряда ферментов, АТФ, присоединяясь к их регуляторным центрам, усиливает или подавляет их активность.
АТФ является также непосредственным предшественником синтеза циклического аденозинмонофосфата — вторичного посредника передачи в клетку гормонального сигнала.
Также известна роль АТФ в качестве медиатора в синапсах.
[Править]Пути синтеза
В организме АТФ синтезируется путём фосфорилирования АДФ:
АДФ + H3PO4 + энергия → АТФ + H2O.
Фосфорилирование АДФ возможно двумя способами: субстратное фосфорилирование и окислительное фосфорилирование (используя энергию окисляющихся веществ). Основная масса АТФ образуется на мембранах митохондрий в ходе окислительного фосфорилирования H-зависимой АТФ-синтазой. Субстратное фосфорилирование АТФ не требует участия мембранных ферментов, оно происходит в процессе гликолиза или путём переноса фосфатной группы с других макроэргических соединений.
Реакции фосфорилирования АДФ и последующего использования АТФ в качестве источника энергии образуют циклический процесс, составляющий суть энергетического обмена.
В организме АТФ является одним из самых часто обновляемых веществ, так у человека продолжительность жизни одной молекулы АТФ менее 1 мин. В течение суток одна молекула АТФ проходит в среднем 2000—3000 циклов ресинтеза (человеческий организм синтезирует около 40 кг АТФ в день), то есть запаса АТФ в организме практически не создаётся, и для нормальной жизнедеятельности необходимо постоянно синтезировать новые молекулы АТФ.
Циклический аденозинмонофосфат (циклический AMФ, цAMФ, cAMP) — производное АТФ, выполняющее в организме роль вторичного посредника, использующегося для внутриклеточного распространения сигналов некоторых гормонов (например,глюкагона или адреналина), которые не могут проходить через клеточную мембрану. Метаболизм цAMФ
цAMФ синтезируется аденилатциклазой в ответ на некоторые гормональные стимуляторы; действует как вторичный посредник при клеточном гормональном контроле путем стимуляции протеинкиназ. цАМФ является аллостерическим эффектором протеинкиназ A и ионных каналов. Синтезируется цАМФ мембранными аденилатциклазами (семейство ферментов, катализирующих реакцию циклизации АТФ с образованием цАМФ и неорганического пирофосфата). Расщепление цАМФ с образованием АМФ катализируется фосфодиэстеразами. Ингибируются цАМФ только при высоких концентрациях метилированных производных ксантина, например, кофеина. Аденилатциклазы активируются G-белками (активность которых в свою очередь зависит от метаботропных рецепторов, связанных с G-белками) .