

55 Гликолиз
Гликолиз — это расщепление глюкозы до молочной кислоты в анаэробных условиях. Гликолиз состоит из двух стадий: подготовительной и главной. эробным гликолизом называют процесс окисления глюкозы до пировиноградной кислоты, протекающий в присутствии кислорода. Все ферменты, катализирующие реакции этого процесса, локализованы в цитозоле клетки. В аэробном гликолизе можно выделить 2 этапа.
1. Подготовительный этап, в ходе которого глюкоза фосфорилируется и расщепляется на две молекулы фосфотриоз. Эта серия реакций протекает с использованием 2 молекул АТФ.
2. Этап, сопряжённый с синтезом АТФ. В результате этой серии реакций фосфотриозы превращаются в пируват. Энергия, высвобождающаяся на этом этапе, используется для синтеза 10 моль АТФ.
Реакции аэробного гликолиза:
1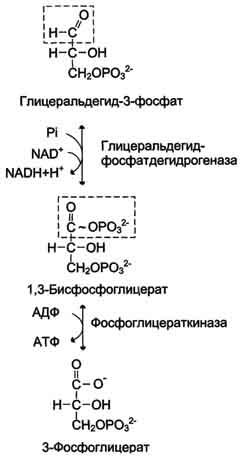
 )
Превращение глюкозо-6-фосфата в 2 молекулы
глицеральдегид-3-фосфата.
Глюкозо-6-фосфат, образованный в результате
фосфорилирования глюкозы с участием
АТФ, в ходе следующей реакции превращается
в фруктозо-6-фосфат. Эта обратимая реакция
изомеризации протекает под действием
фермента глюкозофосфатизомеразы. Затем
следует ещё одна реакция фосфорилирования
с использованием фосфатного остатка и
энергии АТФ. В ходе этой реакции,
катализируемой фосфофруктокиназой,
фруктозо-6-фосфат превращается в
фруктозо-1,6-бисфосфат. Данная реакция,
так же, как гексокиназная, практически
необратима, и, кроме того, она наиболее
медленная из всех реакций гликолиза.
Реакция, катализируемая фосфофруктокиназой,
определяет скорость всего гликолиза,
поэтому, регулируя активность
фосфофруктокиназы, можно изменять
скорость катаболизма глюкозы.
Фруктозо-1,6-бисфосфат далее расщепляется
на 2 триозофосфата: глицеральдегид-3-фосфат
и дигидроксиацетонфосфат. Реакцию
катализирует фермент фруктозобисфосфатальдолаза,
или просто альдолаза. Этот фермент
катализирует как реакцию альдольного
расщепления, так и альдольной конденсации,
т.е. обратимую реакцию. Продукты реакции
альдольного расщепления - изомеры. В
последующих реакциях гликолиза
используется только глицеральдегид-3-фосфат,
поэтому дигидроксиацетонфосфат
превращается с участием фермента
триозофосфатизомеразы в
глицероальдегид-3-фосфат.
)
Превращение глюкозо-6-фосфата в 2 молекулы
глицеральдегид-3-фосфата.
Глюкозо-6-фосфат, образованный в результате
фосфорилирования глюкозы с участием
АТФ, в ходе следующей реакции превращается
в фруктозо-6-фосфат. Эта обратимая реакция
изомеризации протекает под действием
фермента глюкозофосфатизомеразы. Затем
следует ещё одна реакция фосфорилирования
с использованием фосфатного остатка и
энергии АТФ. В ходе этой реакции,
катализируемой фосфофруктокиназой,
фруктозо-6-фосфат превращается в
фруктозо-1,6-бисфосфат. Данная реакция,
так же, как гексокиназная, практически
необратима, и, кроме того, она наиболее
медленная из всех реакций гликолиза.
Реакция, катализируемая фосфофруктокиназой,
определяет скорость всего гликолиза,
поэтому, регулируя активность
фосфофруктокиназы, можно изменять
скорость катаболизма глюкозы.
Фруктозо-1,6-бисфосфат далее расщепляется
на 2 триозофосфата: глицеральдегид-3-фосфат
и дигидроксиацетонфосфат. Реакцию
катализирует фермент фруктозобисфосфатальдолаза,
или просто альдолаза. Этот фермент
катализирует как реакцию альдольного
расщепления, так и альдольной конденсации,
т.е. обратимую реакцию. Продукты реакции
альдольного расщепления - изомеры. В
последующих реакциях гликолиза
используется только глицеральдегид-3-фосфат,
поэтому дигидроксиацетонфосфат
превращается с участием фермента
триозофосфатизомеразы в
глицероальдегид-3-фосфат.
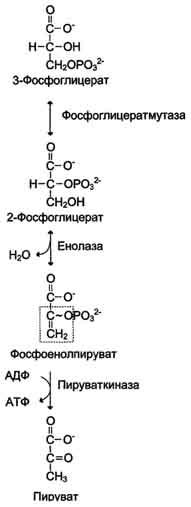
2) Превращение глицеральдегид-3-фосфата в пируват. Эта часть аэробного гликолиза включает реакции, связанные с синтезом АТФ. Наиболее сложной в данной серии реакций является реакция превращения глицеральдегид-3-фосфата в 1,3-бисфосфоглицерат. Это превращение - первая реакция окисления в ходе гликолиза. Реакцию катализирует глицеральдегид-3-фосфатдегидрогеназа, которая является NAD-зависимым ферментом. Значение данной реакции заключается не только в том, что образуется восстановленный кофермент, окисление которого в дыхательной цепи сопряжено с синтезом АТФ, но также и в том, что свободная Е окисления концентрируется в макроэргической связи продукта реакции. Глицеральдегид- 3 -фосфатдегидрогеназа содержит в активном центре остаток цистеина, сульфгидрильная группа которого принимает непосредственное участие в катализе. Окисление глицеральдегид-3-фосфата приводит к восстановлению NAD и образованию с участием Н3РО4 высокоэнергетической ангидридной связи в 1,3-бисфосфоглицерате в положении 1. В следующей реакции высокоэнергетический фосфат передаётся на АДФ с образованием АТФ. Фермент, катализирующий это превращение, назван по обратной реакции фосфоглицераткиназой. Образование АТФ описанным способом не связано с дыхательной цепью, и его называют субстратным фосфорилированием АДФ. Образованный 3-фосфоглицерат уже не содержит макроэргической связи. В следующих реакциях происходят внутримолекулярные перестройки, смысл которых сводится к тому, что низкоэнергетический фосфоэфир переходит в соединение, содержащее высокоэнергетический фосфат. Внутримолекулярные преобразования заключаются в переносе фосфатного остатка из положения 3 в фосфоглицерате в положение 2. Затем от образовавшегося 2-фосфоглицерата отщепляется молекула воды при участии фермента енолазы. В результате реакции образуется замещённый енол - фосфоенолпируват. Образованный фосфоенолпируват - макроэргическое соединение, фосфатная группа которого переносится в следующей реакции на АДФ при участии пируваткиназы. Превращение фосфоенолпирувата в пируват - необратимая реакция. Это вторая в ходе гликолиза реакция субстратного фосфорилирования. Образующаяся енольная форма пирувата затем неферментативно переходит в более термодинамически стабильную кетоформу.
NADH, образующийся при окислении глицеральдегид-3-фосфата в аэробном гликолизе, подвергается окислению путём переноса атомов водорода в митохондриальную дыхательную цепь. Так как сама молекула НАДН через мембрану не проходит, то существуют специальные системы, принимающие атомы водорода от НАДН в цитоплазме и отдающие их в матриксе митохондрий. Эти системы получили название челночные системы. Определены две основные челночные системы – глицеролфосфатная и малат-аспартатная. Ключевыми ферментами глицеролфосфатного челнока являются изоферменты глицерол-3-фосфат-дегидрогеназы – цитоплазматический и митохондриальный. Они отличаются своими коферментами: у цитоплазматической формы – НАД, у митохондриальной – ФАД. В цитозоле метаболиты гликолиза – диоксиацетонфосфат и НАДН образуют глицерол-3-фосфат, поступающий в матрикс митохондрий. Там он окисляется с образованием ФАДН2. Далее ФАДН2 направляется в дыхательную цепь и используется для получения энергии. Таким образом, в результате действий челнока цитозольный НАДН+H+ как бы "превращается" в митохондриальный ФАДН2. Этот челнок активен в печени и белых скелетных мышцах и необходим для получения энергии из глюкозы при работе клетки. Ключевыми ферментами малат-аспартатного челнока являются изоферменты малатдегидрогеназы – цитоплазматический и митохондриальный. Он является распространенным по всем тканям. Этот механизм более сложен: постоянно идущие в цитоплазме реакции трансаминирования аспарагиновой кислоты поставляют оксалоацетат, который под действием цитозольного пула малатдегидрогеназы и за счет "гликолитического" НАДН восстанавливается до яблочной кислоты (малата). Последняя антипортом с α-кетоглутаратом проникает в митохондрии и, являясь метаболитом ЦТК, окисляется в оксалоацетат с образованием НАДН. Так как мембрана митохондрий непроницаема для оксалоацетата, то он аминируется до аспарагиновой кислоты, которая в обмен на глутамат выходит в цитозоль. Таким образом, атомы водорода от цитозольного НАДН перемещаются в состав митохондриального НАДН.
Выход АТФ. Если в клетке имеется кислород, то НАДН из гликолиза направляется в митохондрию (челночные системы), на процессы окислительного фосфорилирования, и там его окисление приносит дивиденды в виде трех молекул АТФ. Обр-ся в гликолизе пируват в аэробных условиях превращается в ПВК-‑дегидрогеназном комплексе в ацетил-S-КоА, при этом образуется 1 молекула НАДН. Ацетил-S-КоА вовлекается в ЦТК и, окисляясь, дает 3 молекулы НАДН, 1 молекулу ФАДН2, 1 молекулу ГТФ. Молекулы НАДН и ФАДН2 движутся в дыхательную цепь (посмотреть), где при их окислении в сумме образуется 11 молекул АТФ. В целом при сгорании одной ацетогруппы в ЦТК образуется 12 молекул АТФ. Суммируя результаты окисления "гликолитического" и "пируватдегидрогеназного" НАДН, "гликолитический" АТФ, энергетический выход ЦТК и умножая все на 2, получаем 38 молекул АТФ. Суммарное уравнение: С6Н12О6 + 6О2 → 6СО2 + Н2О + 2880 кДж/моль.
Аэробный распад глюкозы происходит во многих органах и тканях и служит основным, хотя и не единственным, источником энергии для жизнедеятельности.
. Анаэробный распад глюкозы: последовательность реакций, физиологиче¬ское значение. Роль анаэробного распада глюкозы в мышцах. Дальнейшая судьба молочной кислоты.
В анаэробном процессе пировиноградная кислота восстанавливается до молочной кислоты (лактата), поэтому в микробиологии анаэробный гликолиз называют молочнокислым брожением. Лактат является метаболическим тупиком и далее ни во что не превращается, единственная возможность утилизовать лактат – это окислить его обратно в пируват. Многие клетки организма способны к анаэробному окислению глюкозы. Для эритроцитов он является единственным источником энергии. Клетки скелетной мускулатуры за счет бескислородного расщепления глюкозы способны выполнять мощную, быструю, интенсивную работу, как, например, бег на короткие дистанции, напряжение в силовых видах спорта. Бескислородное окисление глюкозы усиливается при гипоксии клеток при анемиях, нарушении кровообращения в тканях. Гликолиз. Анаэробное превращение глюкозы локализуется в цитозоле и включает два этапа из 11 ферментативных реакций.
Первый этап гликолиза – подготовительный, здесь происходит затрата энергии АТФ, активация глюкозы и образование из нее триозофосфатов.
Первая реакция гликолиза сводится к превращению глюкозы в реакционно-способное соединение за счет фосфорилирования 6-го, не включенного в кольцо, атома углерода. Эта реакция является первой в любом превращении глюкозы, катализируется гексокиназой.
Вторая реакция необходима для выведения еще одного атома углерода из кольца для его последующего фосфорилирования (фермент изомераза). В результате образуется фруктозо-6-фосфат.
Третья реакция – фермент фосфофруктокиназа фосфорилирует фруктозо-6-фосфат с образованием почти симметричной молекулы фруктозо-1,6-дифосфата. Эта реакция является главной в регуляции скорости гликолиза.
В четвертой реакции фруктозо-1,6-дифосфат разрезается пополам фруктозо-1,6-дифосфат-альдолазой с образованием двух фосфорилированных триоз-изомеров – альдозы глицеральдегида (ГАФ) и кетозы диоксиацетона (ДАФ).
Пятая реакция – переход глицеральдегидфосфата и диоксиацетонфосфата друг в друга при участии триозофосфатизомеразы. Равновесие реакции сдвинуто в пользу диоксиацетонфосфата, его доля составляет 97%, доля глицеральдегидфосфата – 3%. Эта реакция, при всей ее простоте, определяет дальнейшую судьбу глюкозы:
при нехватке энергии в клетке и активации окисления глюкозы диоксиацетонфосфат превращается в глицеральдегидфосфат, который далее окисляется на втором этапе гликолиза, при достаточном количестве АТФ, наоборот, глицеральдегидфосфат изомеризуется в диоксиацетонфосфат, и последний отправляется на синтез жиров.
Второй этап гликолиза – это освобождение энергии, содержащейся в глицеральдегидфосфате, и запасание ее в форме АТФ.
Шестая реакция гликолиза (фермент глицеральдегидфосфат-дегидрогеназа) – окисление глицеральдегидфосфата и присоединение к нему фосфорной кислоты приводит к образованию макроэргического соединения 1,3-дифосфоглицериновой кислоты и НАДН.
В седьмой реакции (фермент фосфоглицераткиназа) энергия фосфоэфирной связи, заключенная в 1,3-дифосфоглицерате тратится на образование АТФ. Реакция получила дополнительное название – реакция субстратного фосфорилирования, что уточняет источник энергии для получения макроэргической связи в АТФ (от субстрата реакции) в отличие от окислительного фосфорилирования (от электрохимического градиента ионов водорода на мембране митохондрий).
Восьмая реакция – синтезированный в предыдущей реакции 3-фосфоглицерат под влиянием фосфоглицератмутазы изомеризуется в 2-фосфоглицерат.
Девятая реакция – фермент енолаза отрывает молекулу воды от 2-фосфоглицериновой кислоты и приводит к образованию макроэргической фосфоэфирной связи в составе фосфоенолпирувата.
Д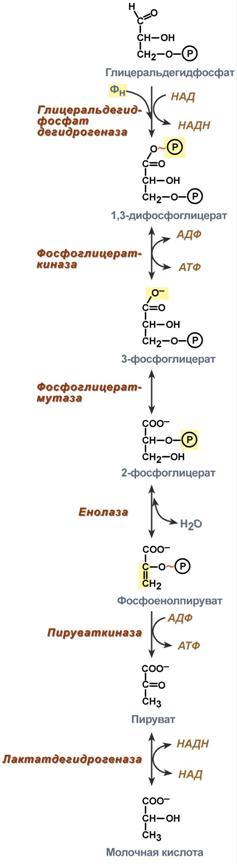 есятая
реакция
гликолиза – еще одна реакция субстратного
фосфорилирования – заключается в
переносе пируваткиназой макроэргического
фосфата с фосфоенолпирувата на АДФ и
образовании пировиноградной кислоты.
есятая
реакция
гликолиза – еще одна реакция субстратного
фосфорилирования – заключается в
переносе пируваткиназой макроэргического
фосфата с фосфоенолпирувата на АДФ и
образовании пировиноградной кислоты.
Р![]() еакция
одиннадцатая
– образование молочной кислоты из
пирувата под действием лактатдегидрогеназы.
Важно то, что эта реакция осуществляется
только в анаэробных условиях. Эта реакция
необходима клетке, так как НАДН,
образующийся в 6-й реакции, в отсутствие
кислорода не может окисляться в
митохондриях.
еакция
одиннадцатая
– образование молочной кислоты из
пирувата под действием лактатдегидрогеназы.
Важно то, что эта реакция осуществляется
только в анаэробных условиях. Эта реакция
необходима клетке, так как НАДН,
образующийся в 6-й реакции, в отсутствие
кислорода не может окисляться в
митохондриях.
При наличии кислорода пировиноградная кислота переходит в митохондрию и превращается в ацетил-S-КоА.
Для стабильного протекания гликолиза в бескислородных условиях необходимы ряд простых условий:-поступление глюкозы, -наличие минимальных количеств АТФ для активации процесса на первом этапе и АДФ как акцептора для фосфатов на втором этапе, -пополнение запасов окисленного НАД для шестой реакции. Если первые два условия подразумеваются сами собой, то для решения третьего условия существует специальное решение: в анаэробных условиях образуемый в шестой, глицеральдегидфосфат-дегидрогеназной реакции, НАДН используется в одиннадцатой реакции для восстановления пирувата до лактата. Образуемый таким образом НАД опять возвращается в шестую реакцию. Следовательно, клетке не нужны дополнительные источники НАД и дополнительные способы уборки НАДН! Процесс поддерживается самостоятельно. Процесс циклического восстановления и окисления НАД в реакциях анаэробного окисления глюкозы получил название гликолитическая оксидоредукция. В аэробных условиях гликолитической оксидоредукции не происходит, НАДН отдает свои атомы водорода на челночные системы для их передачи в дыхательную цепь митохондрий.
Для расчета количества АТФ, образованной при окислении глюкозы необходимо учитывать:
1. Реакции, идущие с затратой или образованием АТФ и ГТФ,
2. Реакции, продуцирующие НАДН и ФАДН2 и использующие их,
3. Так как глюкоза образует две триозы, то все соединения, образующиеся ниже ГАФ-дегидрогеназной реакции, образуются в двойном (относительно глюкозы) количестве.
Расчет АТФ при анаэробном окислении. На подготовительном этапе на активацию глюкозы затрачивается 2 молекулы АТФ, фосфат каждой из которых оказывается на триозе – глицеральдегидфосфате и диоксиацетонфосфате. В следующий второй этап входят две молекулы глицеральдегидфосфата, каждая из которых окисляется до пирувата с образованием 2-х молекул АТФ в седьмой и десятой реакциях – реакциях субстратного фосфорилирования. Таким образом, суммируя, получаем, что на пути от глюкозы до пирувата в чистом виде образуется 2 молекулы АТФ. Однако надо иметь в виду и пятую, глицеральдегидфосфат-дегидрогеназную, реакцию, из которой выходит НАДН. Если условия анаэробные, то он используется в лактатдегидрогеназной реакции, где окисляется для образования лактата и в получении АТФ не участвует.
В анаэробных условиях большое количество пировиноградной кислоты превращается в молочную кислоту, которая легко диффундирует во внеклеточное пространство и даже внутрь некоторых менее активных клеток. Следовательно, молочная кислота представляет собой разновидность «водосточного колодца», в котором исчезают конечные продукты гликолиза, что позволяет гликолизу осуществляться дольше, чем это могло быть при отсутствии молочной кислоты.Большое количество молочной кислоты, которое образуется во время анаэробного гликолиза, не теряется организмом, т.к. если кислород вновь становится доступным, молочная кислота может подвергнуться обратному превращению в глюкозу или напрямую использоваться для получения энергии. Большая часть этих превращений осуществляется в печени, но в небольших количествах может происходить и в других тканях. Если кислород вновь становится доступным после периода анаэробного метаболизма, молочная кислота быстро превращается в пировиноградную кислоту, НАД-Н и Н+. Большие количества этих веществ немедленно окисляются, образуя значительное количество АТФ. Избыток АТФ может явиться причиной того, что более 75% пировиноградной кислоты вновь превращается в глюкозу. Сердечная мышца обладает способностью превращать молочную кислоту в пировиноградную и затем использовать последнюю для получения энергии. В большинстве случаев это происходит при больших физических нагрузках, когда в кровь поступают значительное количество молочной кислоты из скелетных мышц и суммарно дает существенное количество энергии сердечной мышце.
Эффект Пастера. Это снижение потребления глюкозы и прекращение продукции молочной кислоты клеткой в присутствии кислорода. Биохимический механизм эффекта заключается в конкуренции за пируват между пируватдегидрогеназой, превращающей пируват в ацетил-S-КоА, и лактатдегидрогеназой, превращающей пируват в лактат.
У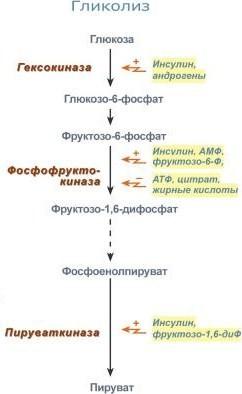 пируватдегидрогеназы сродство гораздо
выше и в обычных аэробных условиях она
окисляет большую часть пировиноградной
кислоты. Как только поступление кислорода
уменьшается (недостаток кровообращения,
тромбоз и т.п.) происходит следующее:
пируватдегидрогеназы сродство гораздо
выше и в обычных аэробных условиях она
окисляет большую часть пировиноградной
кислоты. Как только поступление кислорода
уменьшается (недостаток кровообращения,
тромбоз и т.п.) происходит следующее:
-внутримитохондриальные процессы дыхания не идут и НАДН в дыхательной цепи не окисляется,
-моментально накапливающийся в митохондриях НАДН тормозит цикл трикарбоновых кислот,
-ацетил-S-КоА не входит в ЦТК и ингибирует ПВК-дегидрогеназу.
В этой ситуации пировиноградной кислоте не остается ничего иного как превращаться в молочную.
При наличии кислорода ингибирование ПВК-дегидрогеназы прекращается и она, обладая большим сродством к пирувату, выигрывает конкуренцию.
Регуляция гликолиза
Гликолиз стимулируется инсулином, повышающим количество молекул гексокиназы, фосфофруктокиназы, пируваткиназы.
В печени активность глюкокиназы регулируется гормонами: активацию вызывает инсулин и андрогены, подавляют ее активность глюкокортикоиды и эстрогены.
Для метаболической регуляции чувствительной является фосфофруктокиназа. Она активируется АМФ и собственным субстратом, ингибируется – АТФ, лимонной кислотой, жирными кислотами. Пируваткиназа активируется фруктозо-1,6-дифосфатом. Гексокиназа непеченочных клеток ингибируется продуктом собственной реакции – глюкозо-6-фосфатом.
Основным способом получения АТФ в клетке является окислительное фосфорилирование, протекающее в структурах внутренней мембраны митохондрий. При этом энергия атомов водорода молекул НАДН и ФАДН2, образованных в гликолизе, ЦТК, окислении жирных кислот, преобразуется в энергию связей АТФ. Однако также есть другой способ фосфорилирования АДФ до АТФ – субстратное фосфорилирование, поскольку он является частью метаболического пути («субстратной цепи»). Их особенностью является катализирование растворимыми ферментами. Этот способ связан с передачей макроэргического фосфата или энергии макроэргической связи какого-либо вещества (субстрата) на АДФ. К таким веществам относятся метаболиты гликолиза (1,3-дифосфоглицериновая кислота, фосфоенолпируват), цикла трикарбоновых кислот (сукцинил-SКоА) и креатинфосфат. Энергия гидролиза их макроэргической связи выше, чем 7,3 ккал/моль в АТФ, и роль указанных веществ сводится к использованию этой энергии для фосфорилирования молекулы АДФ до АТФ. Отличия: разные источники энергии, для окислительного необходимы движение электронов в дых цепи, для субстратного необ-ма энергия макроэргической связи.
56 Пентозофосфатный путь
Пентозофосфатный путь превращения глюкозы. Окислительный путь образования пентоз. Представление о неокислительном пути образования гексоз. Распространение, роль, регуляция.
Глюкоза – это субстрат для получения энергии. Энергетика любой клетки нашего организма основана на окислении глюкозы. Окисление глюкозы происходит по двум направлениям:
1.Окисление с образованием пентоз: рибозы, рибулозы, ксилулозы. Этот путь называется пентозофосфатный шунт и не связан с получением Е.
2. Окисление с получением Е.
Так как в кл непрерывно просходят реакции синтеза белков, то для этого процесса требуются рибонуклеиновые кислоты. В свою очередь для синтеза самих нуклеиновых кислот, а точнее пуриновых и пиримидиновых нуклеотидов, требуется рибозо-5-фосфат. Если клетка готовится к делению, то для синтеза ДНК ей нужны дезоксирибонуклеотиды, которые образуются при участии НАДФН. Молекулы НАДФН также используются:
-для синтеза жирных к-т (печень, жировая ткань),
-для синтеза холестерола и др стероидов (печень),
-для синтеза глутаминовой к-ты из α-кетоглутаровой к-ты (реакция восстановительного аминирования),
-для систем антиоксидантной защиты клетки от свободно-радикального окисления (эритроциты).
В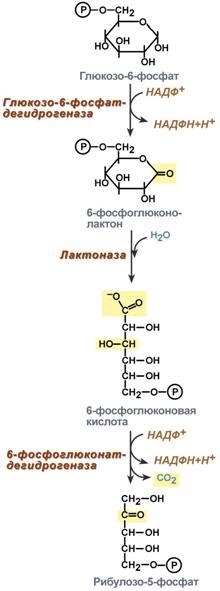 клетке существует процесс, обеспечивающий
одновременное образование рибозы и
НАДФН – это пентозофосфатный
путь.
Наиболее активно реакции пентозофосфатного
пути идут в цитозоле клеток печени,
жировой ткани, эритроцитах, коре
надпочечников, молочной железе при
лактации, в гораздо меньшей степени в
скелетных мышцах. Этот путь окисления
глюкозы не связан с образованием энергии,
а обеспечивает анаболизм клеток. В связи
с этим у новорожденных и детей первых
лет жизни его активность довольно
высока. Пентозофосфатный путь включает
два этапа – окислительный и структурных
перестроек (неокислительный).
клетке существует процесс, обеспечивающий
одновременное образование рибозы и
НАДФН – это пентозофосфатный
путь.
Наиболее активно реакции пентозофосфатного
пути идут в цитозоле клеток печени,
жировой ткани, эритроцитах, коре
надпочечников, молочной железе при
лактации, в гораздо меньшей степени в
скелетных мышцах. Этот путь окисления
глюкозы не связан с образованием энергии,
а обеспечивает анаболизм клеток. В связи
с этим у новорожденных и детей первых
лет жизни его активность довольно
высока. Пентозофосфатный путь включает
два этапа – окислительный и структурных
перестроек (неокислительный).
На первом, окислительном, этапе глюкозо-6-фосфат в трех реакциях превращается в рибулозо-5-фосфат, реакции сопровождаются восстановлением двух молекул НАДФ до НАДФН.
Н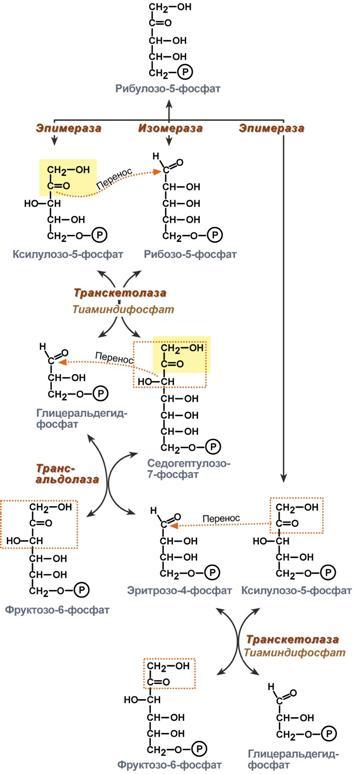 а
этом этапе происходит регуляция процесса:
инсулин повышает активность
глюкозо-6-фосфат-дегидрогеназы и
фосфоглюконат-дегидрогеназы.
а
этом этапе происходит регуляция процесса:
инсулин повышает активность
глюкозо-6-фосфат-дегидрогеназы и
фосфоглюконат-дегидрогеназы.
В окислительной части пентозофосфатного пути глюкозо-6-фосфат подвергается окислительному декарбоксилированию, в результате которого образуются пентозы. Этот этап включает 2 реакции дегидрирования. Суммарное уравнение: Глюкозо-6-фосфат + 2 NADP+ + Н2О → Рибулозо-5-фосфат + 2 NADPH + Н+ + СО2.
Реакции окислительного пути протекают только в том случае, если восстановленный ко-фермент NADPH возвращается в исходное окисленное состояние NADP+ при участии NADPH-зависимых дегидрогеназ. Если потребности клетки в NADPH незначительны, рибо-зо-5-фосфат образуется в результате обратимых реакций неокислительного этапа пентозофосфатного пути, используя в качестве исходных в-в метаболиты гликолиза - глицеральдегид-3-фосфат и фруктозо-6-фосфат.
Роль: Реакции окислительного этапа служат основным источником NADPH в клетках. Гидрированные коферменты снабжают водородом биосинтетические процессы, окислительно-восстановительные реакции, включающие защиту клеток от активных форм О2.
Второй этап – этап структурных перестроек, благодаря которым пентозы способны возвращаться в фонд гексоз. В этих реакциях рибулозо-5-фосфат изомеризуется до рибозо-5-фосфата и ксилулозо-5-фосфата. Далее под влиянием ферментов транскетолазы и трансальдолазы происходят структурные перестройки с образованием других моносахаридов. При реализации всех реакций второго этапа пентозы превращаются во фруктозо-6-фосфат и глицеральдегидфосфат. Глицеральдегид-3-фосфат в зависимости от условий и вида клеток может либо "проваливаться" во 2-й этап гликолиза либо через диоксиацетонфосфат восстанавливаться до глицерол-3-фосфата и далее направляться в синтез фосфатидной кислоты и далее триацилглицеролов. При необходимости из него могут образоваться и гексозы.
Сумма: 2 Фруктозо-6-фосфат + Глицеральдегид-3-фосфат → 2 Ксилулозо-5-фосфат + Рибозо-5-фосфат 2 Ксилулозо-5-фосфат → 2 Рибулозо-5-фосфат 2 Рибулозо-5-фосфат → 2 Рибозо-5-фосфат.
Т.о. неокислительный путь можно представить как процесс возвращения пентоз в фонд гексоз.
Особенности пентозофосфатного пути в разных клетках.
Если клетка нуждается в больших количествах НАДФН (например, синтез жирных кислот в печени или синтез холестерола), то активно будут идти как 1-й, так и 2-й этапы. Образованные фруктозо-6-фосфат и глицеральдегидфосфат в реакциях глюконеогенеза превратятся в глюкозо-6-фосфат, и цикл начнется вновь.
Если клетке нужны НАДФН и энергия АТФ (как в эритроците), то фруктозо-6-фосфат и глицеральдегидфосфат на выходе из 2-го этапа "провалятся" в гликолитические реакции.
Если клетка растет и делится, то ей необходимы НАДФН и рибозо-5-фосфат. В этом случае 2-й этап идти не будет, весь образуемый на 1-м этапе рибулозо-5-фосфат превратится в рибозо-5-фосфат, который используется для синтеза нуклеотидов. НАДФН будет расходоваться на синтез дезоксирибонуклеотидов.
Пентозофосфатный путь обеспечивает клетки рибозой для синтеза пуриновых и пиримидиновых нуклеотидов и гидрированным ко-ферментом NADPH, который используется в восстановительных процессах. Суммарное уравнение пентозофосфатного пути выражается следующим образом:
3 Глюкозо-6-фосфат + 6 NADP+ → 3 СО2 + 6 (NADPH + Н+) + 2 Фруктозо-6-фосфат + Глицеральдегид-3-фосфат.
Ферменты пентозофосфатного пути, так же, как и ферменты гликолиза, локализованы в цитозоле.