

70 Мобилизация жиров из жировой ткани
Синтезируясь во время и сразу после приема пищи (липогенез) и запасаясь в жировой ткани, триацилглицеролы являются формой хранения насыщенных и мононенасыщенных жирных кислот. Распад триацилглицеролов (триглицеридов) по-другому называется липолиз или мобилизация жира. Он идет в жировых клетках постоянно и обычно существует равновесие между синтезом и распадом ТАГ.
Даже в состоянии покоя организма печень, сердце, скелетные мышцы и другие ткани (кроме эритроцитов и нейроцитов) более 50% энергии получают из окисления жирных кислот, поступающих из жировой ткани благодаря фоновому липолизу. По мере уменьшения резервов глюкозы клетки все больше энергии получают из окисления жирных кислот. Таким образом, насыщенные жирные кислоты выполняют роль своеобразного буфера энергии в организме. Мобилизация триацилглицеролов и окисление жирных кислот активируется при нормальных физиологических стрессовых ситуациях – эмоциональный стресс, мышечная работа, голодание, при патологических состояниях – сахарный диабет I типа, другие гормональные заболевания (гиперкортицизм, гипертиреоз). В результате липолиза в адипоцитах образуются свободный глицерол и жирные кислоты. Глицерол с кровью доставляется в печень и почки, здесь фосфорилируется и окисляется в метаболит гликолиза диоксиацетонфосфат. В зависимости от условий ДАФ может включаться в реакции глюконеогенеза (при голодании, мышечной нагрузке) или окисляться в гликолизе до пировиноградной кислоты. Жирные кислоты транспортируются в крови в комплексе с альбуминами плазмы:
-при физической нагрузке – в мышцы,
-в обычных условиях и при голодании – в мышцы и большинство тканей, однако при этом около 30% жирных кислот захватывается печенью.
При голодании и физической нагрузке после проникновения в клетки жирные кислоты вступают на путь β-окисления.
В целом мобилизацию жира можно представить как последовательность следующих событий:
1.Липолиз – гормонзависимый распад ТАГ в жировой ткани или резервных ТАГ в самой клетке.
2.Транспорт жирных кислот из жировой ткани по крови в комплексе с альбумином.
3.Проникновение жирной кислоты в цитозоль клетки-мишени.
4.Активация жирной кислоты через присоединение HS-КоА.
5.Карнитин-зависимое перемещение жирной кислоты в митохондрию.
6.Окисление жирной кислоты с образованием ацетильных групп (в форме ацетил-S-КоА).
7.Сгорание ацетил-S-КоА в цикле лимонной кислоты или синтез (только в печени) кетоновых тел.
В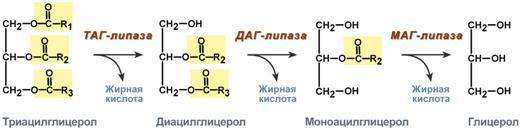 качестве источника энергии могут
использоваться только свободные, т.е.
неэстерифицированные, жирные кислоты.
Поэтому триглицериды сначала гидролизуются
при помощи специфических тканевых
ферментов – липаз – до глицерина и
свободных жирных кислот. Последние из
жировых депо могут переходить в плазму
крови (мобилизация высших жирных кислот),
после чего они используются тканями и
органами тела в качестве энергетического
материала. В жировой ткани содержится
несколько липаз, из которых наибольшее
значение имеют триглицеридлипаза (так
называемая гормоночувствитель-ная
липаза), диглицеридлипаза и
моноглицеридлипаза. Активность двух
последних ферментов в 10–100 раз превышает
активность первого. Три-глицеридлипаза
активируется рядом гормонов (например,
адреналином, норадреналином, глюкагоном
и др.), тогда как диглицеридлипаза и
мо-ноглицеридлипаза не чувствительны
к их действию. Триглицеридлипаза является
регуляторным ферментом. Первое, что
происходит при использовании нейтрального
жира во время голодания и физической
нагрузки – это активация фермента,
отвечающего за отщепление первой жирной
кислоты от триацилглицерола. Фермент
называется гормон-чувствительная
триацилглицерол-липаза или ТАГ-липаза.
Кроме ТАГ-липазы, в адипоцитах имеются
еще диацилглицерол-липаза ( ДАГ-липаза)
и моноацилглицерол-липаза (МАГ-липаза),
активность которых высока и постоянна,
однако в покое эта активность не
проявляется из-за отсутствия субстрата.
Как только в клетке после работы
ТАГ-липазы появляются диацилглицеролы
начинает работать постоянно активная
ДАГ-липаза, продукт ее реакции
моноацилглицерол (МАГ) является субстратом
для МАГ-липазы. Образующиеся жирные
кислоты и глицерол покидают клетку. Для
регуляции активности ТАГ-липазы
обязательно наличие гормонального
влияния (адреналин, глюкагон, соматотропин,
инсулин и др).
качестве источника энергии могут
использоваться только свободные, т.е.
неэстерифицированные, жирные кислоты.
Поэтому триглицериды сначала гидролизуются
при помощи специфических тканевых
ферментов – липаз – до глицерина и
свободных жирных кислот. Последние из
жировых депо могут переходить в плазму
крови (мобилизация высших жирных кислот),
после чего они используются тканями и
органами тела в качестве энергетического
материала. В жировой ткани содержится
несколько липаз, из которых наибольшее
значение имеют триглицеридлипаза (так
называемая гормоночувствитель-ная
липаза), диглицеридлипаза и
моноглицеридлипаза. Активность двух
последних ферментов в 10–100 раз превышает
активность первого. Три-глицеридлипаза
активируется рядом гормонов (например,
адреналином, норадреналином, глюкагоном
и др.), тогда как диглицеридлипаза и
мо-ноглицеридлипаза не чувствительны
к их действию. Триглицеридлипаза является
регуляторным ферментом. Первое, что
происходит при использовании нейтрального
жира во время голодания и физической
нагрузки – это активация фермента,
отвечающего за отщепление первой жирной
кислоты от триацилглицерола. Фермент
называется гормон-чувствительная
триацилглицерол-липаза или ТАГ-липаза.
Кроме ТАГ-липазы, в адипоцитах имеются
еще диацилглицерол-липаза ( ДАГ-липаза)
и моноацилглицерол-липаза (МАГ-липаза),
активность которых высока и постоянна,
однако в покое эта активность не
проявляется из-за отсутствия субстрата.
Как только в клетке после работы
ТАГ-липазы появляются диацилглицеролы
начинает работать постоянно активная
ДАГ-липаза, продукт ее реакции
моноацилглицерол (МАГ) является субстратом
для МАГ-липазы. Образующиеся жирные
кислоты и глицерол покидают клетку. Для
регуляции активности ТАГ-липазы
обязательно наличие гормонального
влияния (адреналин, глюкагон, соматотропин,
инсулин и др).
В целом последовательность событий активации липолиза выглядит следующим образом:
1.Молекула гормона (адреналин, глюкагон, АКТГ) взаимодействует со своим рецептором.
2.Активный гормон-рецепторный комплекс воздействует на мембранный G-белок.
3.G-белок активирует фермент аденилатциклазу.
4.Аденилатциклаза превращает АТФ в цАМФ – вторичный посредник (мессенджер).
5. цАМФ аллостерически активирует фермент протеинкиназу А.
6.Протеинкиназа А фосфорилирует ТАГ-липазу и активирует ее.
7.ТАГ-липаза
отщепляет от триацилглицеролов жирную
кислоту в 1 или 3 положении с образованием
диацилглицерола (ДАГ).А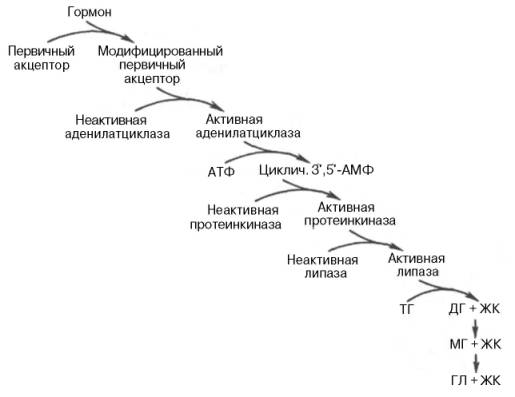 ктивная
триглицеридлипаза расщепляет триглицерид
на диглицерид и жирную кислоту. Затем
при действии ди- и моноглицеридлипаз
образуются конечные продукты липо-лиза
– глицерин и свободные жирные кислоты,
которые поступают в кровяное русло.
ктивная
триглицеридлипаза расщепляет триглицерид
на диглицерид и жирную кислоту. Затем
при действии ди- и моноглицеридлипаз
образуются конечные продукты липо-лиза
– глицерин и свободные жирные кислоты,
которые поступают в кровяное русло.
Скорость липолиза триглицеридов не является постоянной, она подвержена регулирующему влиянию различных факторов, среди которых особое значение имеют нейрогормональные.
Связанные с альбуминами плазмы крови в виде комплекса свободные жирные кислоты с током крови попадают в органы и ткани, где комплекс распадается, а жирные кислоты подвергаются либо β-окислению, либо частично используются для синтеза триглицеридов, глицерофосфолипидов, сфингофосфолипидов и других соединений, а также на эстерификацию холестерина. При физической работе и других состояниях организма, требующих повышенных энергозатрат, потребление триглицеридов жировой ткани как энергетического резерва увеличивается.
Активируют: катехоламины (адреналин), глюкагон, тироксин, кортизол, цАМФ. Ингибирует: инсулин.
Л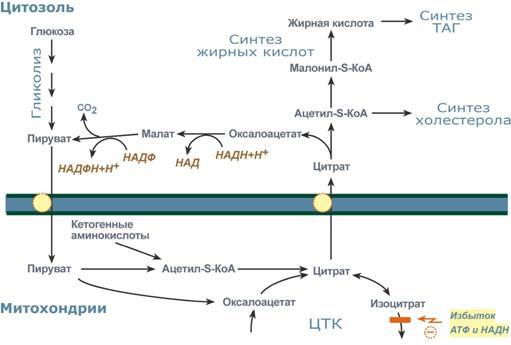 ипогенез
– это синтез жира из глицерина и жирных
к-т. Происходит в печени и жировой ткани.
Глицерин и жирные к-ты должны быть в
активной форме. В жировой тк глицеролкиназа
отсутствует, поэтому активная форма
обр-ся из ДОАФ – метаболита гликолиза
(из глюкозы). Синтез жира протекает в
адбсортивный период (после еды). Молекулы
жира в адипоцитах объединяются крупной
каплей и явл-ся компактной формой
хранения Е запасов. Обмен жира в жировой
тк протекает очень активно: обновляется
полностью за несколько дней. В печени
ежесуточно обр-ся 20-50г жира, который
поступает в кровь в составе ЛПОНП.
Ключевой фермент: ацилтрансфераза.
Активируют: инсулин, эстрогены и АТФ.
Ингибируют: катехоламины, СТГ, йодтиронины,
АКТГ, АДФ. Синтез
нейтральных жиров
происходит за счет этерификации
глицеро-1-фосфата двумя активированными
жирными кислотами. Фосфатная группа
образованной фосфатидной кислоты
отщепляется фосфатазами, что приводит
к образованию диацилглицерина, который
далее реагирует с другой активированной
жирной кислотой, образуя триацилглицерин.
Триацилглицерины составляют энергетическое
депо организма. Они обладают очень
высокой теплотой окисления, равной 37,6
кДж/моль. Жиры локализованы в жировых
клетках (адипоцитах) и характеризуются
высокой скоростью метаболизма. Их
превращение непосредственно регулируется
гормонами, в частности инсулином и
адреналином. Реакции биосинтеза
липидов
могут идти в гладкой эндоплазматической
сети клеток всех органов. Субстратом
для синтеза жиров de novo является глюкоза.
Как известно, попадая в кл, глюкоза
превращается в гликоген, пентозы и
окисляется до пировиноградной кислоты.
При высоком поступлении глюкоза
используется для синтеза гликогена, но
этот вариант ограничивается объемом
клетки. Поэтому глюкоза "проваливается"
в гликолиз и превращается в пируват
либо напрямую, либо через пентозофосфатный
шунт. Во втором случае образуется НАДФН,
который понадобится впоследствии для
синтеза жирных к-т. Пируват переходит
в митохондрии и декарбоксилируется в
ацетил-SКоА и вступает в ЦТК. Однако в
состоянии покоя, при отдыхе, при наличии
избыточного количества энергии в клетке
реакции ЦТК (в частности,
изоцитратдегидрогеназная реакция)
блокируются избытком АТФ и НАДН. В
результате накапливается первый
метаболит ЦТК – цитрат. По градиенту
концентрации он перемещается в цитозоль,
расщепляется с образованием ацетил-SКоА,
который далее используется в биосинтезе
холестерола, жирных кислот и
триацилглицеролов. Оксалоацетат, также
образуемый из цитрата, восстанавливается
до яблочной кислоты и возвращается в
митохондрии *посредством малат-аспартатного
челнока (на рисунке не показан), *после
декарбоксилирования малата до пирувата
НАДФ-зависимым малик-ферментом.
Образованный НАДФН будет использован
при синтезе жирных кислот.
ипогенез
– это синтез жира из глицерина и жирных
к-т. Происходит в печени и жировой ткани.
Глицерин и жирные к-ты должны быть в
активной форме. В жировой тк глицеролкиназа
отсутствует, поэтому активная форма
обр-ся из ДОАФ – метаболита гликолиза
(из глюкозы). Синтез жира протекает в
адбсортивный период (после еды). Молекулы
жира в адипоцитах объединяются крупной
каплей и явл-ся компактной формой
хранения Е запасов. Обмен жира в жировой
тк протекает очень активно: обновляется
полностью за несколько дней. В печени
ежесуточно обр-ся 20-50г жира, который
поступает в кровь в составе ЛПОНП.
Ключевой фермент: ацилтрансфераза.
Активируют: инсулин, эстрогены и АТФ.
Ингибируют: катехоламины, СТГ, йодтиронины,
АКТГ, АДФ. Синтез
нейтральных жиров
происходит за счет этерификации
глицеро-1-фосфата двумя активированными
жирными кислотами. Фосфатная группа
образованной фосфатидной кислоты
отщепляется фосфатазами, что приводит
к образованию диацилглицерина, который
далее реагирует с другой активированной
жирной кислотой, образуя триацилглицерин.
Триацилглицерины составляют энергетическое
депо организма. Они обладают очень
высокой теплотой окисления, равной 37,6
кДж/моль. Жиры локализованы в жировых
клетках (адипоцитах) и характеризуются
высокой скоростью метаболизма. Их
превращение непосредственно регулируется
гормонами, в частности инсулином и
адреналином. Реакции биосинтеза
липидов
могут идти в гладкой эндоплазматической
сети клеток всех органов. Субстратом
для синтеза жиров de novo является глюкоза.
Как известно, попадая в кл, глюкоза
превращается в гликоген, пентозы и
окисляется до пировиноградной кислоты.
При высоком поступлении глюкоза
используется для синтеза гликогена, но
этот вариант ограничивается объемом
клетки. Поэтому глюкоза "проваливается"
в гликолиз и превращается в пируват
либо напрямую, либо через пентозофосфатный
шунт. Во втором случае образуется НАДФН,
который понадобится впоследствии для
синтеза жирных к-т. Пируват переходит
в митохондрии и декарбоксилируется в
ацетил-SКоА и вступает в ЦТК. Однако в
состоянии покоя, при отдыхе, при наличии
избыточного количества энергии в клетке
реакции ЦТК (в частности,
изоцитратдегидрогеназная реакция)
блокируются избытком АТФ и НАДН. В
результате накапливается первый
метаболит ЦТК – цитрат. По градиенту
концентрации он перемещается в цитозоль,
расщепляется с образованием ацетил-SКоА,
который далее используется в биосинтезе
холестерола, жирных кислот и
триацилглицеролов. Оксалоацетат, также
образуемый из цитрата, восстанавливается
до яблочной кислоты и возвращается в
митохондрии *посредством малат-аспартатного
челнока (на рисунке не показан), *после
декарбоксилирования малата до пирувата
НАДФ-зависимым малик-ферментом.
Образованный НАДФН будет использован
при синтезе жирных кислот.
Какой процесс будет преобладать в организме - синтез жиров (липогенез) или их распад (липолиз), зависит от поступления пищи и физической активности. В абсорбтивном состоянии под действием инсулина происходит липогенез, в постабсорбтивном состоянии - липолиз, активируемый глюкагоном. Адреналин, секреция которого увеличивается при физической активности, также стимулирует липолиз.
глюкагон в печени активируется синтез жиров. В жировой ткани индуцируется синтез ЛП-липазы в адипоцитах и осуществляется её экспонирование на поверхность эндотелия; следовательно, в этот период увеличивается поступление жирных кислот в адипоциты. Одновременно инсулин активирует белки-переносчики глюкозы - ГЛЮТ-4. Поступление глюкозы в адипоциты и гликолиз также активируются. В результате образуются все необходимые компоненты для синтеза жиров: глицерол-3-фосфат и активные формы жирных кислот. В печени инсулин, действуя через различные механизмы, активирует ферменты путём дефосфорилирования и индуцирует их синтез. В результате увеличиваются активность и синтез ферментов, участвующих в превращении части глюкозы, поступающей с пищей, в жиры. Это - регуляторные ферменты гликолиза, пируватдегидрогеназный комплекс и ферменты, участвующие в синтезе жирных кислот из ацетил-КоА. Результат действия инсулина на обмен углеводов и жиров в печени - увеличение синтеза жиров и секреция их в кровь в составе ЛПОНП. ЛПОНП доставляют жиры в капилляры жировой ткани, где действие ЛП-липазы обеспечивает быстрое поступление жирных кислот в адипоциты, где они депонируются в составе триацилглицеринов.
Запасание жиров в жировой ткани - основная форма депонирования источников энергии в организме человека (табл. 8-6). Запасы жиров в организме человека массой 70 кг составляют 10 кг, но у многих людей количество жиров может быть значительно больше.
Жиры образуют в адипоцитах жировые вакуоли. Жировые вакуоли иногда заполняют значительную часть цитоплазмы. Скорость синтеза и мобилизации подкожного жира происходит неравномерно в разных частях организма, что связано с неодинаковым распределением рецепторов гормонов на адипоцитах.
Регуляция мобилизации жиров. Мобилизация депонированных жиров стимулируется глюкагоном и адреналином и, в меньшей степени, некоторыми другими гормонами (соматотроп-ным, кортизолом). В постабсорбтивный период и при голодании глюкагон, действуя на адипоциты через аденилатциклазную систему, активирует протеинкиназу А, которая фосфо-рилирует и, таким образом, активирует гормончувствительную липазу, что инициирует липо-лиз и выделение жирных кислот и глицерина в кровь. При физической активности увеличивается секреция адреналина, который действует через β-адренергические рецепторы адипоцитов, активирующие аденилатциклазную систему (рис. 8-24). В настоящее время обнаружено 3 типа β-рецепторов: β1, β2, β3, активация которых приводит к липолитическому действию. К наибольшему липолитическому действию приводит активация β3-рецепторов. Адреналин одновременно действует и на α2-рецепторы адипоцитов, связанные с ингибирующим G-белком, что инактивирует аденилатциклазную систему. Вероятно, действие адреналина двояко: при низких концентрациях в крови преобладает его антилиполитическое действие через α2-рецепторы, а при высокой - преобладает липолитическое действие через β-рецепторы.
Для мышц, сердца, почек, печени при голодании или физической работе жирные кислоты становятся важным источником энергии. Печень перерабатывает часть жирных кислот в кетоновые тела, используемые мозгом, нервной тканью и некоторыми другими тканями как источники энергии.
В результате мобилизации жиров концентрация жирных кислот в крови увеличивается приблизительно в 2 раза (рис. 8-25), однако абсолютная концентрация жирных кислот в крови невелика даже в этот период. Т1/2 жирных кислот в крови тоже очень мал (менее 5 мин), что означает существование быстрого потока жирных кислот из жировой ткани к другим органам. Когда постабсорбтивный период сменяется аборбтивным, инсулин активирует специфическую фосфатазу, которая дефосфорилирует гормончувствительную липазу, и распад жиров останавливается.