

Природа гена. Эволюция представлений о гене.
Ген (от. греч. Genos – род, происхождение) - дискретная единица наследственности, участок молекулы нуклеиновой кислоты, который характеризуется специфической последовательностью нуклеотидов и представляет собой единицу функции, отличной от функций других генов, способный изменяться мутированием.
Геномом называется одинарный полный набор генетического материала организма. В него входят последовательности нуклеотидов ДНК гаплоидного набора хромосом, ДНК митохондрии и хлоропластов.
У гена есть структура и функции.
Структура – это то, из чего состоит и как организован ген, функции – что и как он делает
Термин «ген» предложил датский ученый В. Иогансен 1909г.
Г.Мендель ввел понятие «Наследственный фактор»
На основе точных экспериментов сделал обобщения относительно свойств и поведения их при передачи от родителей потомкам. Эти обобщения легли в основу теории гена.
Свойства наследственных факторов следующие:
Наличие альтернативных наследственных факторов для развития каждого конкретного признака организма (в современном представлении доминантный и рецессивный аллели гена).
Парность наследственных факторов, определяющих развитие признака (у диплоидного организма). Существенный вывод: наследуются не признаки, а передаются от родителей к потомкам вместе с гаметами гены.
Дискретность и относительное постоянство гена. В гибридной зиготе рецессивный аллель не сливается и не смешивается с доминантным аллелем, а поступает в гамету F1 в чистом виде и, объединяясь с подобным аллелем при оплодотворении, проявляется как рецессивный признак в F2 . Этот феномен в последующем получил название закон чистоты гамет.
Наследственный фактор к началу двадцатого века выступал как условная единица наследственности.
Дальнейшая конкретизация представлений о гене связана с работами школы американского ученого Т.Моргана.
Даказательства локализации генов в хромосоме:
Гены, наследующиеся сцеплено с полом (локализация генов в половых хромосомах –Х или Y)
Сцепленное наследование группы признаков в отличии от правил независимого наследования признаков Г.Менделя. Было показано наличие определенного числа групп сцепление генов, соответственно гаплоидному числу хромосом конкретного биологического вида.
Обмен генами между гомологичным хромосомами (генетические и цитологические доказательства кроссинговера), приводящие к рекомбинации генов.
Величина генетической рекомбинации (процента кроссинговера - перекреста) отражает расстояние между генами одной группы сцепления: чем больше отстоят друг от друга гены, тем больше процент кроссинговера. Гены в хромосоме располагаются в линейном порядке, и каждый ген имеет определенное местоположение – локус.
Итак, к концу 20-х годов ХХ века ген представляли как обособленный участок хромосомы, который контролирует один определенный признак, изменяющийся как единое целое и неделимое при кроссинговере.
В 1929 г. А.С. Серебровский и Н.П. Дубинин экспериментально доказали, что ген не представляет собой единицу мутации, он имеет сложную структуру – состоит из нескольких субъединиц, способных самостоятельно мутировать (ступенчатый аллелизм, или Центровая теория гена).
Эти ученые выявили сложное строение гена у дрозофилы, отвечающего за редукцию щетинок. Задний отдел груди имеет в норме 4 щетинки. Мутации отдельных участков этого сложного гена вызывают отсутствие разных щетинок. Такое явление было названо ступенчатым аллелизмом. На примере различных мутаций этого гена авторами было показано, что ген не всегда мутирует как единое целое: мутации могут затрагивать различные его участки.
Новаторская идея о делимости гена
не сразу получила признание среди
генетиков, однако множество данных,
полученных на различных объектах, в
конце концов убедили генетическое
сообщество в том, что ген не является
единицей мутации и рекомбинации.
Несколько позже идея о сложном строении гена была подкреплена Э. Льюисом, М. Грином и другими учеными, исследовавшими внутригенный кроссинговер на дрозофиле.
Таким образом, ген представляли как участок хромосомы, контролирующий развитие конкретного признака, имеющий определенную линейную протяженность и способный мутировать в разных участках и быть разделенным кроссинговером. Ген комплексен, так как его отдельные участки могут различаться по функциям.
По Менделю 1 ген- 1 признак.
1902год А. Гаррод (английский врач) исследовал
родословные семей с АЛКАПТОНУРИЕЙ. Это болезнь наследуется по аутосомно-
рецессивному типу и связана с нарушением обмена веществ.
Обусловлена врожденной ошибкой метаболизма (пигментация соединительной ткани, артрит, моча черного цвета).
Повреждение одного гена вызывает отсутствие одной биохимической реакции. А так как биохимическая реакция катализируется ферментом то, предположил А.Гаррод, ген предопределяет наличие активного фермента.
В 40х годах XX века Дж. Бидл и Э. Тейтум выдвинули концепцию «1 ген – 1 фермент».
Они использовали новый подход при изучении метаболизма у микроскопического грибка – нейроспоры. Д.Бидл и Э.Тейтум получали мутации, у которых отсутствовала активность того или иного фермента метаболизма. Это приводило к тому, что мутантный гриб был не способен сам синтезировать определенный метаболит и жил только тогда, когда этот метаболит добавляли в питательную среду. Так было сформулировано важнейшее положение биохимической генетики «один ген – один фермент». Это положение означает, что один ген кодирует фермент, катализирующий одну из биохимических реакций.
В последующем она была трансформирована в концепцию «один ген – одна полипептидная цепь».
В синтезе белка гемоглобина участвуют два неаллельных гена. Каждый отвечает за синтез своей полипептидной цепи. В цитоплазме клетки эти цепи объединяются, образуя функциональную структуру гемоглобина – тетрамер.
Белок гемоглобин
2 неаллельных гена
неаллельных гена
ген I ген β

I- полипептидная β - полипептидная
цепь цепь
(141 аминокислота) (146 аминокислот)

 Две
I цепи Две β
цепи
Две
I цепи Две β
цепи
т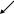 етрамер
гемоглобина
етрамер
гемоглобина
1 ген – 1 – полипептидная цепь
Доказательство роли ДНК в передаче наследственной информации
1.
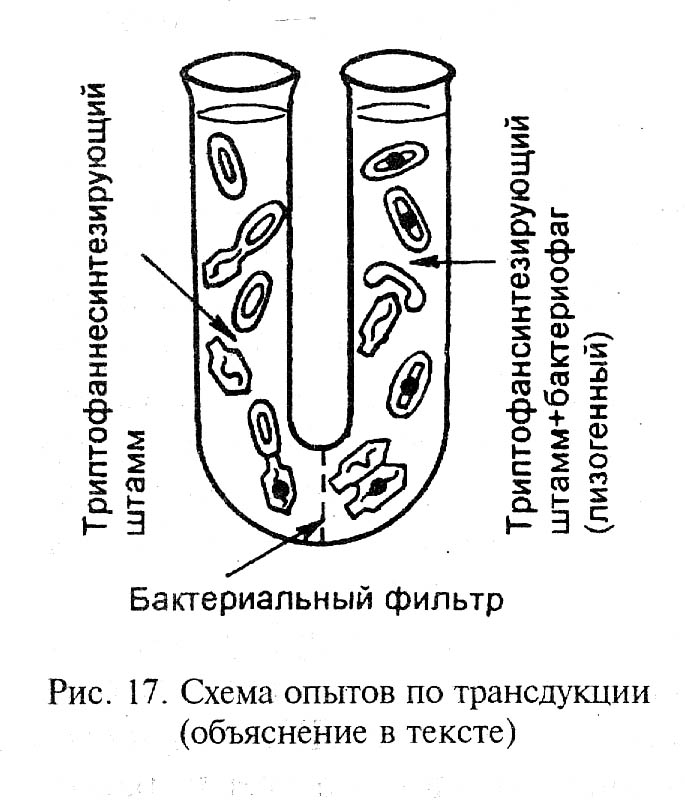
Доказательство роли ДНК в передачи наследственной информации получили Н.Циндер и Дж.Ледерберг. В 1952 г. они описали явление трансдукции. U-образную трубку заполняли жидкой питательной средой и посредине ставили бактериальный фильтр. В левое колено помещали триптофаннесинтезирующий штамм (22А) бактерий мышиного тифа, а в правое – триптофансинтезирующий штамм бактерий дикого типа (2А). В правое колено добавляли бактериофаг (вирус, паразитирующий на бактериях). Через некоторое время в левом колене появлялись триптофансинтезирующие бактерии. Непосредственного контакта между бактериями не было. Роль «переносчика» этого свойства выполнили бактериофаги. Размножаясь в бактериях штамма 2А, они встраивали в свою ДНК частицы ДНК клеток хозяина. Проходя бактериальный фильтр и внедряясь в бактерии штамма 22А, они передавали им участок ДНК, ответственный за синтез триптофана.
Трансдукция – это способность бактериофагов переносить фрагменты ДНК от одного штамма бактерий к другому и передавать соответствующие свойства.
2.

Трансформация – это способность одного штамма бактерий встраивать в свою ДНК участки молекулы ДНК другого штамма и приобретать при этом свойства последнего.
3.
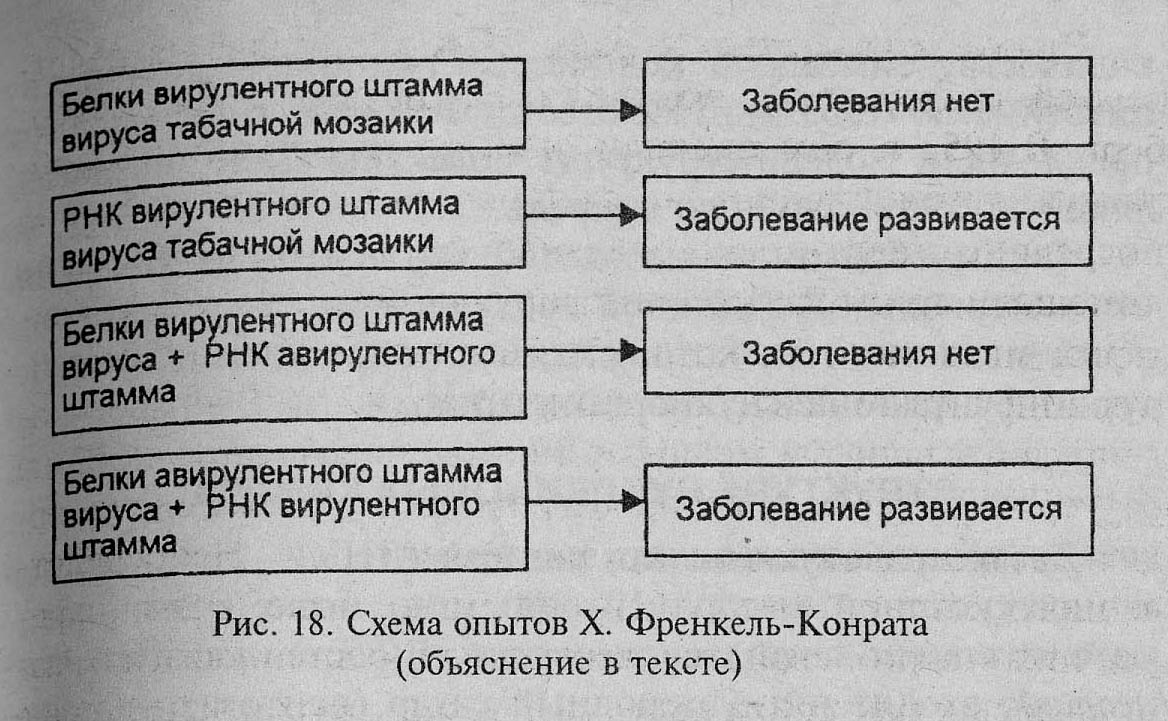
Х.Френкель-Конрат провел опыты (1950г.) с вирусом табачной мозаики (ВТМ). ВТМ состоит из белковой оболочки и РНК. С помощью химических методов удалось разделить вирус на РНК и белок. При введении в листья растений табака белковой части вируса заболевание не развивалось, а при введении РНК на листьях появлялась мозаика. Этот опыт является доказательством того, что нуклеиновые кислоты, а не белки являются носителями генетической информации.
Конец 50-х годов Сеймур Бензер
Ген дискретная единица.
При выполнении основной функции (программирование синтеза белка) ген выступает как целостная единица, изменение которой вызывает перестройку
структуры белковой молекулы.
Эту единицу Бензер назвал цистроном.
Дискретность гена заключается в наличии у него субъединиц.
Единица мутации – мутон.
Единица рекомбинации - рекон
Современные представления о структуре гена, его функционировании, регуляции его активности складывалось во второй половине ХХ века. Важными вехами на этом пути стали:
- открытие двухспиральной структуры ДНК;
- выделение РНК и выяснение ее роли в передаче наследственной информации от ДНК к РНК и белку;
- расшифровка генетического кода.
Ген – это определенный участок молекулы ДНК, состоящий их нескольких тысяч пар нуклеотидов, способных мутировать и быть разделенными рекомбинацией. Однако функционально он представляет единое целое.
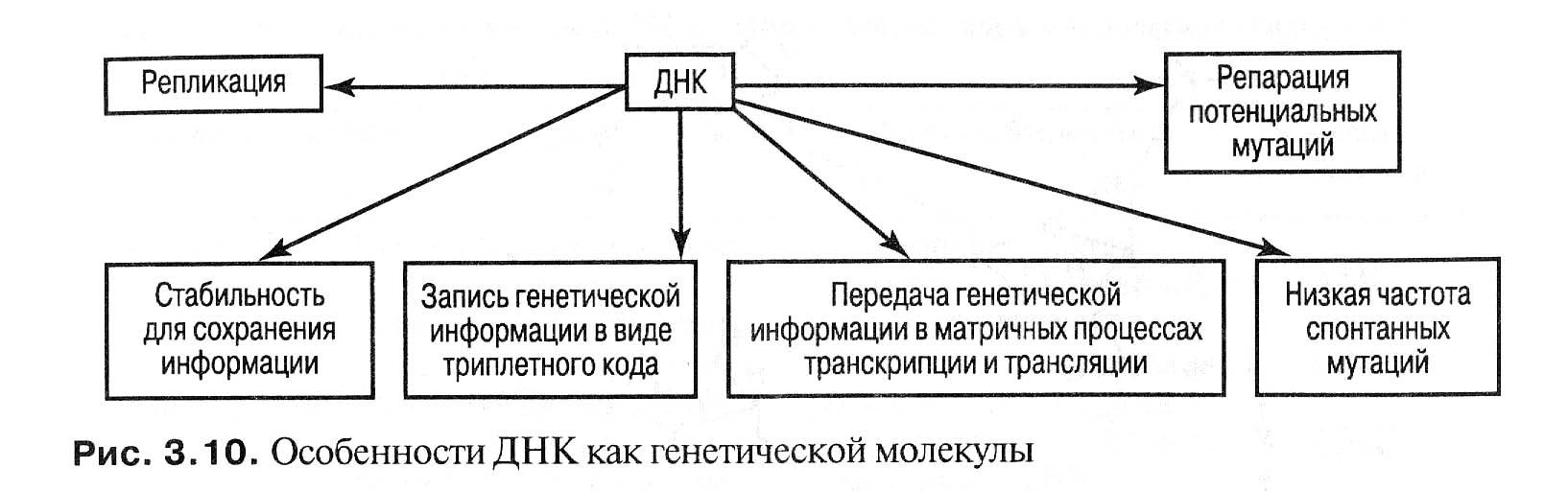
Стабильность. Она обеспечивается водородными, гликозидными и фосфодиэфирными связями, а также механизмом репарации спонтанных и индуцированных повреждений;
Способность к репликации. Благодаря этому механизму в соматических клетках сохраняется диплоидное число хромосом;
Наличие генетического кода. Последовательность оснований в ДНК с помощью процессов транскрипции и трансляции преобразуется в последовательность аминокислот в полипептидной цепи;
Способность к генетической рекомбинации. Благодаря этому механизму образуются новые сочетания сцепленных генов.
Геном вирусов.
Генетический материал вирусов представлен одной молекулой нуклеиновой кислоты (ДНК и РНК), окруженной защитной белковой оболочкой – капсидом.
Вирусы
РНК содержащие ДНК содержащие
~3000-7000 ДНК фага Т4 – 180*10³п.н.
нуклеотидов и кодирует более 40 белков.
Крупные молекулы ДНК компактно упакованы внутри капсида благодаря суперсперилизации.
Вирусы - внутриклеточные паразиты.
Варианты развития вируса в клетке:
Интеграция с геномом хозяина – лизогения.
Синтез вирусных частиц на основе генетической программы вируса, но с помощью метаболической системы хозяина – лизис. Этот вариант приводит к разрушению клетки – хозяина.
Структура гена вирусов.
В
 ирусы
ирусы
Одноцепочечная Двуцепочечная Одноцепочечная Двуцепочечная

 РНК
РНК ДНК ДНК
РНК
РНК ДНК ДНК


(представляют собой
вариант соединенных
цепей без расхождения
после синтеза второй цепи)
линейные и кольцевые
формы ДНК
РНК – содержащие вирусы
с «плюс - цепью» - могут сразу экспрессироваться.
с «минус - цепью» - вначале строят «плюс - цепь» с помощью РНК – полимеразы клетки - хозяина.
Для вирусов характерно явление «перекрывающихся генов» – «ген в гене» (1977г. Ф. Сэнджер)
Такая организация генетического материала позволяет экономно использовать небольшие информационные возможности генома. Однако она имеет свои ограничения, поскольку одна мутация может «выключить» сразу два или более генов.

РНК – вирусы могут встроиться в молекулу ДНК хозяина только с помощью фермента, получившего название обратной транскриптазы. После проникновения вируса в клетку, обратная транскриптаза осуществляет синтез сначала одноцепочечной комплементарной ДНК, а затем по ее матрице – двухцепочечной ДНК-копии. Затем ДНК-копия вирусного РНК-генома встраивается в хромосомную ДНК клетки хозяина, вместе с ней транскрибируется, а затем транслируется с образованием вирусных белков.
Геном прокариот
представлен одной кольцевой молекулой ДНК, формирующей компактную структуру нуклеоида посредством суперспирализации