

Занятие №8. Плазматическая мембрана (пм). Эндоцитоз и экзоцитоз. Межклеточные контакты.
1. Клеточная мембрана. Строение.
В настоящее время наибольшим признанием пользуется жидкостно-мозаичная модель мембраны, предложенная в 1972 году Сингером и Николсоном (Singer, Nicolson). Согласно этой модели мембрана состоит из бислоя липидов, в котором плавают (или закреплены) белковые молекулы, образуя в нём своеобразную мозаику. Мембранные белки могут пронизывать бислой насквозь (интегральный белок), примыкать к бислою (периферический белок) или погружаться в него. Многие белки мембраны являются гликопротеинами, а мембранообразующие липиды - гликолипидами.
1.1. Липиды.
Жирные кислоты, входящие в состав мембранных липидов, содержат четное число углеродных атомов. Среди этих кислот наиболее часто встречаются:
а) Насыщенные |
б) Ненасыщенные |
пальмитиновая С16:0 стеариновая С18:0 лигноцериновая С24:0
|
пальмитоолеиновая С16:1(7) олеиновая С18:1 (9) нервоновая С24:1 (9) линолевая С18:2 (9,12) линоленовая С18:3 (9,12,15) арахидоновая С20:4 (5,8,11,14)
|
Цифры указывают количество углеродных атомов и число двойных связей, а также их положение (в скобках).
Все липиды клеточной мембраны представляют собой амфипатические молекулы, то есть у них ест гидрофильный и гидрофобный концы.
Одним из факторов, влияющих на текучесть мембраны, является холестерол. Плазматические мембраны эукариот содержат довольно большое количество холестерола – приблизительно 1 молекулу на каждую молекулу фосфолипида. Молекулы холестерола ориентируются в бислое таким образом, чтобы их гидроксильные группы примыкали к полярным головам фосфолипидных молекул. При этом их жесткие стероидные кольца частично иммобилизуют участки углеводородных цепей, непосредственно примыкающих к полярным головам. Хотя холестерол делает липидный бислой менее текучим, при его высоких концентрациях (как в ПМ) он предотвращает слипание и кристаллизацию углеводородных цепей. Кроме того, холестерол увеличивает упругость и механическую плотность бислоя. Именно благодаря холестеролу мембрана может менять свою форму в ответ на приложенную к ней силу. Дело в том, что в отличие от фосфолипидов холестерол может быстро перераспределяться между монослоями.
Определенная текучесть мембраны имеет важное биологическое значение. Об этом свидетельствует тот факт, что бактерии, дрожжи и другие пойкилотермные организмы, принимающие температуру окружающей среды, изменяют жирокислотный состав своих ПМ таким образом, чтобы текучесть мембраны оставалась примерно постоянной. Например, при падении температуры начинают синтезироваться жирные кислоты с большим числом цис-двойных связей, для того чтобы предотвратить уменьшение текучести бислоя.
ПМ бактерий обычно имеют фосфолипид только одного типа и не содержат холестерол (механическая прочность обеспечивается клеточной стенкой). В ПМ животных клеток обнаруживаются 4 основных фосфолипида: фосфатидилхолин (лецитин), сфингомиелин, фосфатидилсерин и фосфатидилэтаноламин. Другие фосфолипиды, такие как инозитолфосфолипиды, важны в функциональном отношении (роль в передаче клеточных сигналов), но представлены в относительно малых количествах.
Гликолипиды.
Гликолипиды – олигосахаридсодержащие липиды. Эти молекулы обнаруживаются только в наружной половине бислоя, а их сахарные группы ориентированы к поверхности клетки. Гликолипиды составляют около 5% липидных молекул наружного монослоя. Они сильно различаются у разных видов и даже в разных тканях одного и того же вида. У бактерий и растений почти все гликолипиды – это производные липидов на основе глицерола, например, широко распространенного фосфотидилохолина. В животных клетках гликолипиды построены на основе церамида, каковым является, например, фосфолипид сфингомиелин.
Все гликолипидные молекулы различаются по числу сахарных остатков в их полярных головах. Так, полярные головы нейтральных гликолипидов содержат от 1 до 15 и более нейтральных (незаряженных) сахаров. Одним из простейших нейтральных гликолипидов является галактоцереброзид, полярная голова которого состоит только из одной галактозы; галактоцереброзид – это главный гликолипид миелина (многослойной мембранной оболочки, которая окружает аксон).
Самые сложные из гликолипидов – ганглиозиды – содержат один или более остатков сиаловой кислоты (N-ацетилнейраминовой кислоты, или NANA), которые придают молекулам ганглиозидов отрицательный заряд. Особенно много ганглиозидов в мембранах нервных клеток.
Установлено, что GМ1-ганглиозид действует как поверхностный рецептор для бактериального токсина, образуемого при холере. Холерный токсин связывается с поверхностью и попадает внутрь только тех клеток, у которых в плазматической мембране присутствует GМ1-ганглиозид (к ним, например, относятся эпителиальные клетки кишечника). Проникновение холерного токсина внутрь клетки приводит к долговременному увеличению концентрации внутриклеточного сАМР, которое обуславливает сильный приток Na+ и воды внутрь кишечника.
1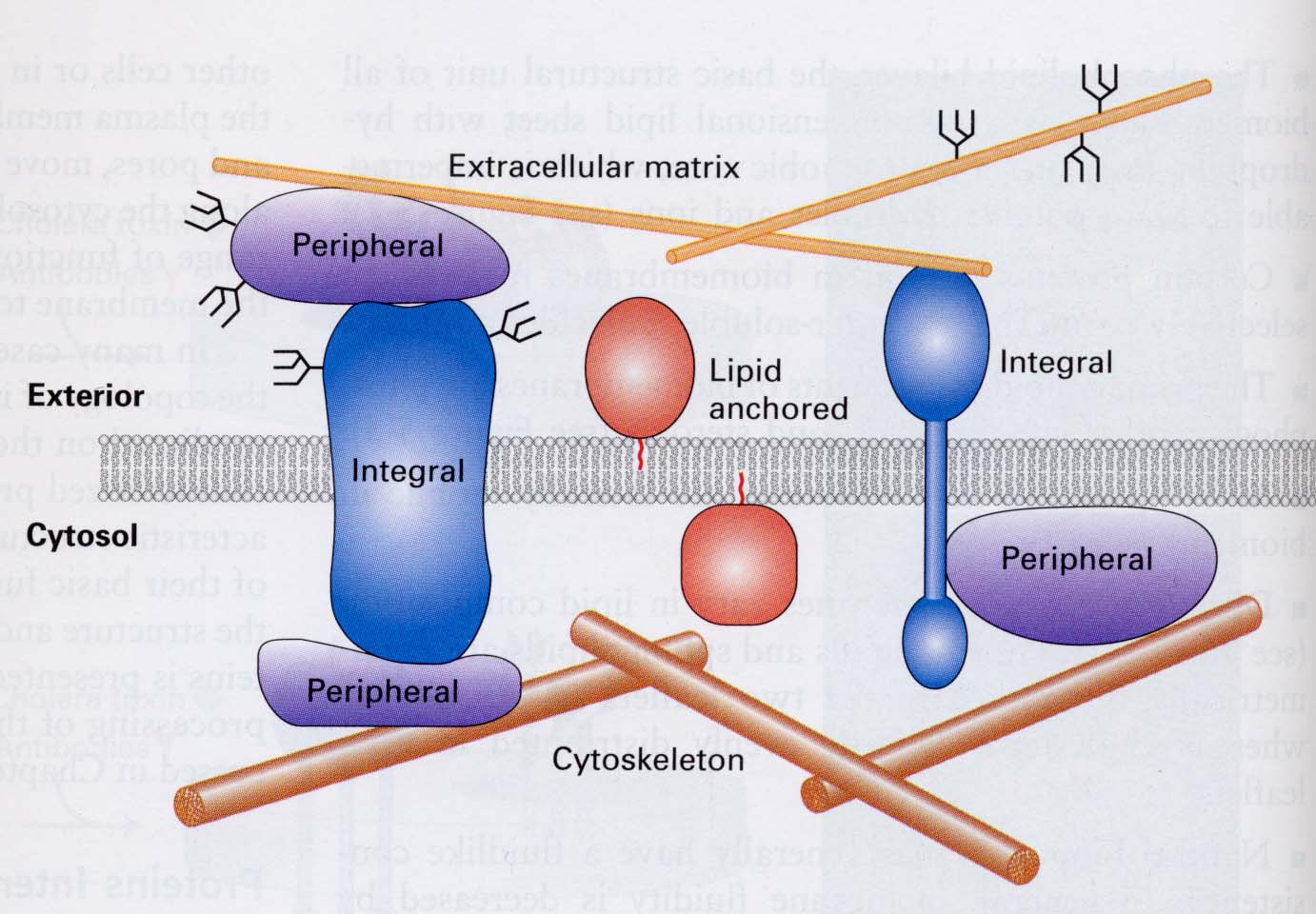 .2.
Белки. Белки
легко встраиваются в фосфолипидный
бислой и удерживаются в нем за счет
гидрофобных взаимодействий неполярных
группировок аминокислот с остатками
жирных кислот. Гидрофильные области
белка образуют электростатические
связи с полярными головками фосфолипидов
и определенным образом ориентированы
в бислое. По типу расположения белка
в фосфолипидном бислое различают
интегральные
и периферические
белки. Участки интегральных белков,
имеющие гидрофобные аминокислоты
погружены в бислой глубоко и могут
пересекать его. Это могут быть
транспортные белки, ферменты, рецепторы
и регуляторные белки. «Заякоривание»
периферических белков происходит с
помощью водородных связей и ионных
взаимодействий. Этому способствует
также наличие в периферическом белке
остатка жирной кислоты или олигосахарида,
которые внедряются в липидный бислой
и служат своеобразным якорем.
.2.
Белки. Белки
легко встраиваются в фосфолипидный
бислой и удерживаются в нем за счет
гидрофобных взаимодействий неполярных
группировок аминокислот с остатками
жирных кислот. Гидрофильные области
белка образуют электростатические
связи с полярными головками фосфолипидов
и определенным образом ориентированы
в бислое. По типу расположения белка
в фосфолипидном бислое различают
интегральные
и периферические
белки. Участки интегральных белков,
имеющие гидрофобные аминокислоты
погружены в бислой глубоко и могут
пересекать его. Это могут быть
транспортные белки, ферменты, рецепторы
и регуляторные белки. «Заякоривание»
периферических белков происходит с
помощью водородных связей и ионных
взаимодействий. Этому способствует
также наличие в периферическом белке
остатка жирной кислоты или олигосахарида,
которые внедряются в липидный бислой
и служат своеобразным якорем.
Многие мембранные белки диффундируют в плоскости мембраны. Мембранные белки так же, как и мембранные липиды, не могут перескакивать с одной стороны бислоя на другую (такой перескок носит название флип-флоп), но они способны вращаться вокруг оси, перпендикулярной плоскости бислоя (вращательная диффузия). К тому же многие мембранные белки могут перемещаться в плоскости мембраны (латеральная диффузия).
Липопротеины.
Липопротеиновые комплексы представляют собой шаровидные агрегаты, состоящие из ядра, образованного неполярными липидами (триацилглицеринами и ацилхолестеринами), и оболочки толщиной примерно 2 нм, построенной из апопротеинов и амфифильных липидов (фосфолипидов и холестерина). Наружная сторона оболочки полярна, вследствие этого липиды растворимы в плазме. Чем больше липидное ядро, т. е. чем большую часть составляют неполярные липиды, тем меньше плотность липопротеинового комплекса.
Липопротеиновые комплексы делятся на пять групп. Ниже они приведены в порядке уменьшения размера и увеличения плотности: это хиломикроны и остатки хиломикронов, липопротеины очень низкой плотности [ЛОНП (VLDL от англ. very low density lipoproteins)], липопротеины промежуточной плотности [ЛПП (IDL от англ. intermediate density lipoproteins)], липопротеины низкой плотности [ЛНП (LOL от англ. low density lipoproteins)], липопротеины высокой плотности [ЛВП (HDL от англ. high density lipoproteins)]. Липопротеиновые комплексы несут на внешней поверхности характерный апопротеин, который «плавает» на оболочке. Апопротеины играют решающую роль в функционировании липопротеинов: они служат молекулами узнавания для мембранных рецепторов (см. ниже) и необходимыми партнерами для ферментов и белков, которые участвуют в метаболизме и обмене липидов.
Хиломикроны обеспечивают транспорт пищевых липидов от кишечника к тканям. Хиломикроны образуются в слизистой кишечника и транспортируются в кровь лимфатической системой. В мышцах и жировой ткани они разрушаются липазой липопротеинов, активирующейся апопротеином С-II. Под действием этого фермента хиломикроны быстро теряют бóльшую часть своих триацилглицеринов. Остатки хиломикронов утилизируются печенью.
ЛОНП, ЛПП и ЛНП тесно связаны между собой. Они транспортируют триацилглицерины, холестерин и фосфолипиды от печени к тканям. ЛОНП образуются в печени и могут превращаться, как и хиломикроны, в ЛПП и ЛНП путем отщепления жирных кислот. Образующиеся ЛНП снабжают холестерином различные ткани организма.
ЛВП возвращают избыточный холестерин, образующийся в тканях, обратно в печень. Во время транспорта холестерин ацилируется жирными кислотами из лецитина. В этом процессе участвует лецитинхолестеринацилтрансфераза. Между ЛВП и ЛОНП также происходит обмен липидами и белками.