

4) Дисахариды
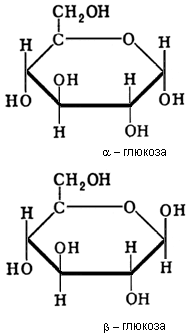
биозы, углеводы, молекулы которых состоят из двух остатков моносахаридов (См. Моносахариды). Все Д. построены по типу гликозидов (См. Гликозиды). При этом водородный атом гликозидного гидроксила одной молекулы моносахарида замещается остатком др. молекулы моносахарида за счёт полуацетального или спиртового гидроксила. В первом случае образуются Д., не обладающие восстанавливающими свойствами, во втором — Д. с восстанавливающими свойствами (I). В группу невосстанавливающих Д. входят трегалоза (микоза, или грибной сахар), состоящая из 2 остатков глюкозы (См. Глюкоза) (II); Сахароза, состоящая из остатков глюкозы и фруктозы (III), и др. К группе восстанавливающих Д. относятся Мальтоза (IV), Целлобиоза (V), Лактоза (VI) и др. Д. могут содержать 5- и 6-членные кольца моносахаридов (пентозы и гексозы) и различаться по конфигурации гликозидной связи (α- или β-гликозиды). Пространственные формы (конформации (См. Конформация)) колец моносахаридных остатков в разных Д. могут варьировать. Так, целлобиоза и мальтоза различаются не только конфигурацией гликозидной связи (α — у мальтозы и β — у целлобиозы), но и тем, что в целлобиозе оба остатка находятся в одинаковой конформации, а в мальтозе — в разных. Д. — хорошо кристаллизующиеся вещества, легко растворимы в воде и в 45 — 48°-ном спирте, плохо растворимы в 96°-ном спирте; оптически активны; сладки на вкус. Гидролиз Д. (для сахарозы называется инверсией) происходит при действии кислот; при наличии 5-членного кольца в моносахаридном остатке скорость кислотного гидролиза Д. возрастает. Гидролиз Д. осуществляется также ферментами (карбогидразами), например α- или β-гликозидазами (в зависимости от типа гликозидной связи в Д.). В результате гидролиза образуются моносахариды.
Д. широко распространены в животных и растительных организмах. Они встречаются в свободном состоянии (как продукты биосинтеза или частичного гидролиза полисахаридов), а также как структурные компоненты гликозидов и др. соединений. Обычно Д. получают из природных источников (например, сахарозу — из сахарной свёклы или сахарного тростника, лактозу — из молока животных). Многие Д. синтезируют химическими и биохимическими методами.
Сахароза, лактоза и мальтоза — ценные пищевые и вкусовые вещества. Производством сахарозы занята сахарная промышленность
5) По своему функциональному назначению гомополисахариды могут быть разделены на две группы: структурные и резервные полисахариды. Важным структурным гомополисахаридом является целлюлоза, а главными резервными – гликоген и крахмал (у животных и растений соответственно).
Строгая
классификация
по химическому строению или биологической
роли вследствие отсутствия для многих
полисахаридов
исчерпывающих данных невозможна. Поэтому
чаще всего полисахариды
«именуются» по источникам выделения,
несмотря на то что один и тот же полисахарид
может быть получен из совершенно разных
источников.Крахмал,
как отмечалось, является основным
резервным материалом растительных
организмов.
В небольших количествах он содержится
в листьях, но главным образом накапливается
в семенах (зерна злаков, например пшеницы,
риса, кукурузы, содержат до 70% крахмала),
а также в луковицах, клубнях и сердцевине
стебля растений, где содержание его
доходит до 30%.Крахмал
представляет собой смесь 2 гомополисахаридов:
линейного – амилозы
и разветвленного – амилопектина,
общая формула которых (С6Н10О5)n.
Как правило, содержание амилозы
в крахмале
составляет 10–30%, амилопектина
– 70–90%. Полисахариды
крахмала
построены из остатков D-глюкозы,
соединенных в амилозе
и линейных цепях амило-пектина
α-1–>4-связями, а в точках ветвления
амилопектина
– межцепочечными α-1–>6-связями:
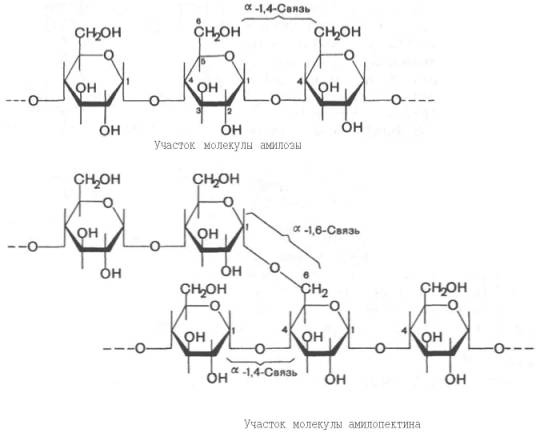
Итак, единственным моносахаридом, входящим в состав крахмала, является D-глюкоза. В молекуле амилозы линейно связано в среднем около 1000 остатков глюкозы; отдельные участки молекулы амилопектина состоят из 20–30 таких единиц. В настоящее время общепринятой является «ветвистая» структура отдельных цепочек с α-1–>4-связями в молекуле амилопектина (рис. 5.3).Известно, что в воде амилоза не дает истинного раствора. Цепочка амилозы в воде образует гидратированные мицеллы. В растворе при добавлении йода амилоза окрашивается в синий цвет. Амилопектин также дает мицеллярный раствор, но форма мицелл несколько иная. Полисахарид амилопектин окрашивается йодом в красно-фиолетовый цвет.Крахмал имеет молекулярную массу 105–107 Да. При частичном кислотном гидролизе крахмала образуются полисахариды меньшей степени полимеризации – декстрины , при полном гидролизе – глюкоза.
Рис. 5.3. Структура крахмала: а - амилоза с характерной для нее спиральной структурой; б - амилопектин, образующий в точках ветвления связи типа 1-6.
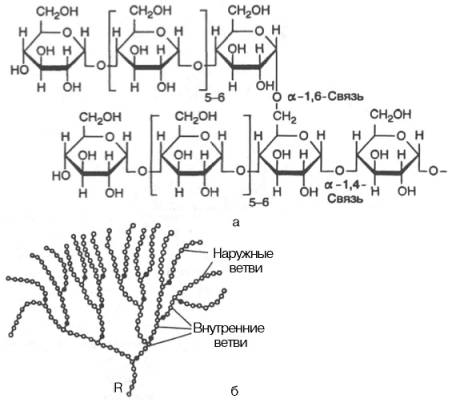
Рис. 5.4. Строение отдельного участка (а) и всей молекулы (б) гликогена (по Майеру). Белые кружки - остатки глюкозы, соединенные α-1,4-связью; черные кружки - остатки глюкозы, присоединенные α-1,6-связью; R - редуцирующая концевая группа. Внутренние цепи, или ветви,- участки между точками ветвления. Наружные цепи, или ветви, начинаются от точки ветвления и кончаются нередуцирующим остатком глюкозы.Для человека крахмал является важным пищевым углеводом; содержание его в муке составляет 75–80%, в картофеле – 25%.Гликоген – главный резервный полисахарид высших животных и человека, построенный из остатков D-глюкозы. Эмпирическая формула гликогена, как и крахмала, (С6Н10О5)n. Гликоген содержится практически во всех органах и тканях животных и человека; наибольшее количество обнаружено в печени и мышцах. Молекулярная масса гликогена 105–108 Да и более. Его молекула построена из ветвящихся полиглюкозидных цепей, в которых остатки глюкозы соединены α-1–>4-гликозидными связями. В точках ветвления имеются α-1–>6-гликозидные связи. По строению гликоген близок к амилопектину. В молекуле гликогена различают внутренние ветви – участки от периферической точки ветвления до нередуци-рующего конца цепи (рис. 5.4).Гликоген характеризуется более разветвленной структурой, чем амило-пектин; линейные отрезки в молекуле гликогена включают 11–18 остатков α-D-глюкопиранозы.При гидролизе гликоген, подобно крахмалу, расщепляется с образованием сначала декстринов, затем мальтозы и, наконец, глюкозы.Инулин – полисахарид, содержащийся в клубнях и корнях георгинов, артишоков и одуванчиков. При его гидролизе образуется фруктоза, следовательно, он представляет собой фруктазан.Метилирование инулина свидетельствует, что остатки D-фруктозы связаны между собой 2–>1-связями и находятся в фуранозной форме:
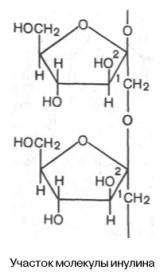
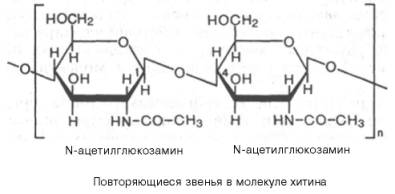
Степень полимеризации инулина равна примерно 35 моносахарным остаткам. Этот полисахарид в отличие от картофельного крахмала легко растворяется в теплой воде. Инулин используют в физиологических исследованиях для определения скорости клубочковой фильтрации в почках.Хитин – важный структурный полисахарид беспозвоночных животных (главным образом членистоногих). Из него, в частности, построен наружный скелет ракообразных и насекомых.Хитин также частично или полностью замещает целлюлозу в клеточных стенках сапрофитных растений, например грибов.Структуру хитина составляют N-ацетил-D-глюкозаминовые звенья, соединенные β-(1–>4)-гликозидными связями:
Целлюлоза (клетчатка) – наиболее широко распространенный структурный полисахарид растительного мира. Он состоит из α-глюкозных остатков в их β-пиранозной форме, т.е. в молекуле целлюлозы β-глюко-пиранозные мономерные единицы линейно соединены между собой β-(1–>4)-связями:
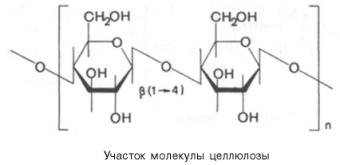
При частичном гидролизе целлюлозы образуется дисахарид целлобиоза, а при полном гидролизе – D-глюкоза. Молекулярная масса целлюлозы 1000–2000 кДа. Клетчатка не переваривается ферментами пищеварительного тракта, так как набор этих ферментов у человека не содержит гидролаз, расщепляющих β-связи. В связи с этим целлюлозу можно рассматривать как значительный неиспользуемый «пищевой» резерв. Вместе с тем известно, что присутствие оптимальных количеств клетчатки в пище способствует формированию кала. При полном исключении клетчатки из пищи нарушается формирование каловых масс.В кишечнике жвачных и других травоядных животных имеются микроорганизмы, способные к ферментативному расщеплению β-связей (β-глю-козидных связей), и для этих животных целлюлоза является важным источником пищевых калорий.Основная масса целлюлозы используется для изготовления хлопчатобумажных тканей и бумаги. Кроме того, на основе целлюлозы производятся искусственные волокна, пластмассы и т.д. Характерной особенностью целлюлозы, определяющей в значительной степени ее механические, физико-химические и химические свойства, является линейная конформация молекул, закрепленная внутримолекулярными водородными связями.
6) Полисахариды, в структуре которых характерно наличие двух или более типов мономерных звеньев, носят название гетерополисахаридов.Принято считать, что, поскольку гетерополисахариды чаще состоят только из двух различных мономеров, расположенных повторяющимся образом, они не являются информационными молекулами [Бохински Р., 1987].Важнейшие представители гетерополисахаридов в органах и тканях животных и человека – гликозаминогликаны (мукополисахариды). Они состоят из цепей сложных углеводов, содержащих аминосахара и уроновые кислоты.
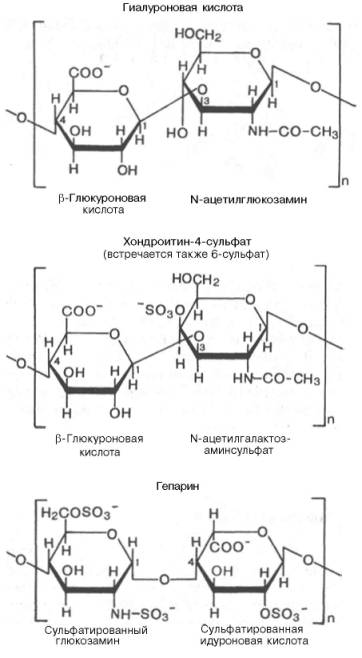
Рис. 5 . 5 . Строение некоторых сложных полисахаридов (гликозамино-гликанов).
Рис. 5.6. Протеогликановый агрегат (схема).
Единая длинная молекула гиалуро-ната (1) нековалентно связана со многими молекулами белка (2), каждая из которых содержит кова-лентно связанные молекулы хонд-роитинсульфата (3) и кератансуль-фата (4).Различают шесть основных классов гликозаминогликанов (см. главу 21). Каждый из гликозаминогликанов содержит характерную для него повторяющуюся дисахаридную единицу; во всех случаях (кроме кератансульфатов) эта единица содержит либо глюкуроновую, либо идуроновую кислоту. Все гликозаминогликаны, за исключением гиалуроновой кислоты, содержат остатки моносахаридов с О- или N-сульфатной группой.Гликозаминогликаны значительно различаются по размерам, их молекулярные массы в пределах от 104 Да для гепарина до 107 Да для гиалуроновой кислоты.Выделенные индивидуальные гликозаминогликаны могут содержать смесь цепей различной длины (рис. 5.5). Гликозаминогликаны как основное скрепляющее вещество связаны со структурными компонентами костей и соединительной ткани. Их функция состоит также в удержании большой массы воды и в заполнении межклеточного пространства. Иными словами, гликозаминогликаны – основной компонент внеклеточного вещества – жела-тинообразного вещества, заполняющего межклеточное пространство тканей. Они также содержатся в больших количествах в синовиальной жидкости – это вязкий материал, окружающий суставы, который служит смазкой и амортизатором.Поскольку водные растворы гликозаминогликанов гелеобразны, их называют мукополисахаридами.Наконец, если цепи гликозаминогликана присоединены к белковой молекуле, соответствующее соединение называют протеогликаном.Протеогликаны образуют основное вещество внеклеточного матрикса. В отличие от простых гликопротеинов, которые содержат только несколько процентов углеводов (по массе), протеогликаны могут содержать до 95% (и более) углеводов (рис. 5.6).
10) Стероиды — вещества животного или реже растительного происхождения, обладающие высокой биологической активностью. Стероиды образуются в природе из изопреноидных предшественников. Особенностью строения стероидов является наличие конденсированной тетрациклической системы гонана (прежнее название — стеран). Ядро гонана в стероидах может быть насыщенным или частично ненасыщенным, содержать алкильные и некоторые функциональные группы — гидроксильные, карбонильные или карбоксильную.[1]В регуляции обмена веществ и некоторых физиологических функций организма участвуют стероидные гормоны. Ряд синтетических гормонов, например, преднизолон, по действию на организм превосходят природные аналоги. В группу стероидов входят содержащиеся в организме человека стероидный спирт холестерин, а также желчные кислоты — соединения, имеющие в боковой цепи карбоксильную группу, например, холевая кислота.К стероидам относятся также сердечные гликозиды — вещества растительного происхождения (из наперстянки, строфанта, ландыша), регулирующие сердечную деятельность. В гликозидах стероидный фрагмент соединен гликозидной связью с олигосахаридом.[2]
11) Аминокисло́ты (аминокарбо́новые кисло́ты) — органические соединения, в молекуле которых одновременно содержатся карбоксильные и аминные группы.Аминокислоты могут рассматриваться как производные карбоновых кислот, в которых один или несколько атомов водорода заменены на аминные группФизические свойстваАминокислоты — бесцветные кристаллические вещества, хорошо растворимые в воде. Многие из них обладают сладким вкусом.
Общие химические свойстваВсе аминокислоты амфотерные соединения, они могут проявлять как кислотные свойства, обусловленные наличием в их молекулах карбоксильной группы —COOH, так и основные свойства, обусловленные аминогруппой —NH2. Аминокислоты взаимодействуют с кислотами и щелочами:
NH2 —CH2 —COOH + HCl → HCl • NH2 —CH2 —COOH (хлороводородная соль глицина)
NH2 —CH2 —COOH + NaOH → H2O + NH2 —CH2 —COONa (натриевая соль глицина)
Растворы аминокислот в воде благодаря этому обладают свойствами буферных растворов, т.е. находятся в состоянии внутренних солей.
NH2 —CH2COOH
![]() N+H3 —CH2COO-
N+H3 —CH2COO-
Аминокислоты обычно могут вступать во все реакции, характерные для карбоновых кислот и аминов.
Этерификация:
NH2 —CH2 —COOH + CH3OH → H2O + NH2 —CH2 —COOCH3 (метиловый эфир глицина)
Важной особенностью аминокислот является их способность к поликонденсации, приводящей к образованию полиамидов, в том числе пептидов, белков, нейлона, капрона.
Реакция образования пептидов:
HOOC —CH2 —NH —H + HOOC —CH2 —NH2 → HOOC —CH2 —NH —CO —CH2 —NH2 + H2O
Изоэлектрической точкой аминокислоты называют значение pH, при котором максимальная доля молекул аминокислоты обладает нулевым зарядом. При таком pH аминокислота наименее подвижна в электрическом поле, и данное свойство можно использовать для разделения аминокислот, а также белков и пептидов.Цвиттер-ионом называют молекулу аминокислоты, в которой аминогруппа представлена в виде -NH3+, а карбоксигруппа — в виде -COO−. Такая молекула обладает значительным дипольным моментом при нулевом суммарном заряде. Именно из таких молекул построены кристаллы большинства аминокислот.Некоторые аминокислоты имеют несколько аминогрупп и карбоксильных групп. Для этих аминокислот трудно говорить о каком-то конкретном цвиттер-ионе.
Получение
Большинство аминокислот можно получить в ходе гидролиза белков или как результат химических реакций:
CH3COOH + Cl2 + (катализатор) → CH2ClCOOH + HCl; CH2ClCOOH + 2NH3 → NH2 —CH2COOH + NH4Cl
Оптическая изомерия
Все входящие в состав живых организмов α-аминокислоты, кроме глицина, содержат асимметричный атом углерода (треонин и изолейцин содержат два асимметричных атома) и обладают оптической активностью. Почти все встречающиеся в природе α-аминокислоты имеют L-форму, и лишь L-аминокислоты включаются в состав белков, синтезируемых на рибосомах.Данную особенность «живых» аминокислот весьма трудно объяснить, так как в реакциях между оптически неактивными веществами L и D-формы образуются в одинаковых количествах. Возможно, выбор одной из форм (L или D) — просто результат случайного стечения обстоятельств: первые молекулы, с которых смог начаться матричный синтез, обладали определенной формой, и именно к ним «приспособились» соответствующие ферменты.
D-аминокислоты в живых организмахАспарагиновые остатки в метаболически неактивных структурных белках претерпевают медленную самопроизвольную неферментативную рацемизацию: так в белках дентина и эмали зубов L-аспартат переходит в D-форму со скоростью ~0,1 % в год[2], что может быть использовано для определения возраста млекопитающих. Рацемизация остатков аспарагиновой также отмечена при старении коллагена, предполагается, что такая рацемизация специфична для аспарагиновой кислоты и протекает за счет образования сукцинимидного кольца при внутремолекулярном ацилировании пептидного азота свободной карбоксильной группой аспарагиновой кислоты[3].С развитием следового аминокислотного анализа D-аминокислоты были обнаружены сначала в составе клеточных стенок некоторых бактерий (1966), а затем и в тканях высших организмов. Так, D-аспартат и D-метионин предположительно являются нейромедиаторами у млекопитающих.В состав некоторых пептидов входят D-аминокислоты, образующиеся при посттрансляционной модификации. Например, D-метионин и D-аланин входят в состав опиоидных гептапептидов кожи южноамериканских амфибий филломедуз (дерморфина, дермэнкефалина и делторфинов). Наличие D-аминокислот определяет высокую биологическую активность этих пептидов как анальгетиков.Сходным образом образуются пептидные антибиотики бактериального происхождения, действующие против грамположительных бактерий — низин, субтилин и эпидермин.Гораздо чаще D-аминокислоты входят в состав пептидов и их производных, образующихся путем нерибосомного синтеза в клетках грибов и бактерий. Видимо, в этом случае исходным материалом для синтеза служат также L-аминокислоты, которые изомеризуются одной из субъединиц ферментного комплекса, осуществляющего синтез пептида.
Протеиногенные аминокислоты
Основная статья: Белки
В процессе биосинтеза белка в полипептидную цепь включаются 20 α-аминокислот, кодируемых генетическим кодом. Помимо этих аминокислот, называемых протеиногенными, или стандартными, в некоторых белках присутствуют специфические нестандартные аминокислоты, возникающие из стандартных в процессе посттрансляционных модификаций. В последнее время к протеиногенным аминокислотам иногда причисляют трансляционно включаемые селеноцистеин (Sec, U) и пирролизин (Pyl, O). Это так называемые 21-я и 22-я аминокислоты.Вопрос, почему именно эти 20 аминокислот стали «избранными», остаётся не решённым. Не совсем ясно, чем эти аминокислоты оказались предпочтительнее других похожих. Например, ключевым промежуточным метаболитом пути биосинтеза треонина, изолейцина и метионина является α-аминокислота гомосерин. Очевидно, что гомосерин — очень древний метаболит, но для треонина, изолейцина и метионина существуют аминоацил-тРНК-синтетазы, тРНК, а для гомосерина — нет.
Структурные формулы 20-ти протеиногенных аминокислот обычно приводят в виде так называемой таблицы протеиногенных аминокислот:
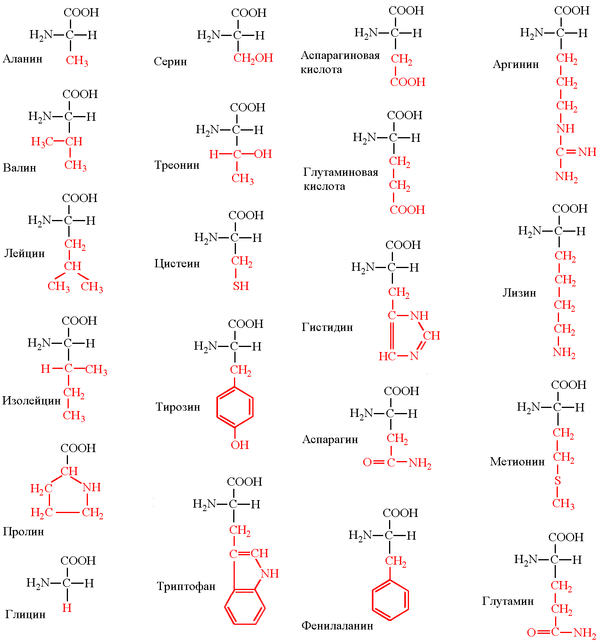
Для запоминания однобуквенного обозначения протеиногенных аминокислот используется мнемоническое правило (последний столбец).
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Классификация
По радикалу
Неполярные: глицин, аланин, валин, изолейцин, лейцин, пролин, метионин, фенилаланин, триптофан
Полярные незаряженные (заряды скомпенсированы) при pH=7: серин, треонин, цистеин, аспарагин, глутамин, тирозин
Полярные заряженные отрицательно при pH<7: аспартат, глутамат
Полярные заряженные положительно при pH>7: лизин, аргинин, гистидин
По функциональным группам
Алифатические
Моноаминомонокарбоновые: глицин, аланин, валин, изолейцин, лейцин
Оксимоноаминокарбоновые: серин, треонин
Моноаминодикарбоновые: аспартат, глутамат, за счёт второй карбоксильной группы несут в растворе отрицательный заряд
Амиды моноаминодикарбоновых: аспарагин, глутамин
Диаминомонокарбоновые: лизин, аргинин, несут в растворе положительный заряд
Серосодержащие: цистеин, метионин
Ароматические: фенилаланин, тирозин, триптофан, (гистидин)
Гетероциклические: триптофан, гистидин, пролин
Иминокислоты: пролин
По классам аминоацил-тРНК-синтетаз
Класс I: валин, изолейцин, лейцин, цистеин, метионин, глутамат, глутамин, аргинин, тирозин, триптофан
Класс II: глицин, аланин, пролин, серин, треонин, аспартат, аспарагин, гистидин, фенилаланин
Для аминокислоты лизин существуют аминоацил-тРНК-синтетазы обоих классов.
По путям биосинтезаПути биосинтеза протеиногенных аминокислот разноплановы. Одна и та же аминокислота может образовываться разными путями. К тому же совершенно различные пути могут иметь очень похожие этапы. Тем не менее, имеют место и оправданы попытки классифицировать аминокислоты по путям их биосинтеза. Существует представление о следующих биосинтетических семействах аминокислот: аспартата, глутамата, серина, пирувата и пентоз. Не всегда конкретную аминокислоту можно однозначно отнести к определённому семейству; делаются поправки для конкретных организмов и учитывая преобладающий путь. По семействам аминокислоты обычно распределяют следующим образом:
Семейство аспартата: аспартат, аспарагин, треонин, изолейцин, метионин, лизин.
Семейство глутамата: глутамат, глутамин, аргинин, пролин.
Семейство пирувата: аланин, валин, лейцин.
Семейство серина: серин, цистеин, глицин.
Семейство пентоз: гистидин, фенилаланин, тирозин, триптофан.
Фенилаланин, тирозин, триптофан иногда выделяют в семейство шикимата.
По способности организма синтезировать из предшественников
Незаменимые
Для большинства животных и человека незаменимыми аминокислотами являются: валин, изолейцин, лейцин, треонин, метионин, лизин, фенилаланин, триптофан, аргинин, гистидин.
Заменимые
Для большинства животных и человека заменимыми аминокислотами являются: глицин, аланин, пролин, серин, цистеин, аспартат, аспарагин, глутамат, глутамин, тирозин.
Классификация аминокислот на заменимые и незаменимые не лишена недостатков. К примеру, тирозин является заменимой аминокислотой только при условии достаточного поступления фенилаланина. Для больных фенилкетонурией тирозин становится незаменимой аминокислотой. Аргинин синтезируется в организме человека и считается заменимой аминокислотой, но в связи с некоторыми особенностями его метаболизма при определённых физиологических состояниях организма может быть приравнен к незаменимым. Гистидин также синтезируется в организме человека, но не всегда в достаточных количествах, потому должен поступать с пищей.
По характеру катаболизма у животных
Биодеградация аминокислот может идти разными путями. По характеру продуктов катаболизма у животных протеиногенные аминокислоты делят на три группы: глюкогенные (при распаде дают метаболиты, не повышающие уровень кетоновых тел, способные относительно легко становиться субстратом для глюконеогенеза: пируват, α-кетоглутарат, сукцинил-KoA, фумарат, оксалоацетат), кетогенные (распадаются до ацетил-KoA и ацетоацетил-KoA, повышающие уровень кетоновых тел в крови животных и человека и преобразующиеся в первую очередь в липиды), глюко-кетогенные (при распаде образуются метаболиты обоих типов).
Глюкогенные: глицин, аланин, валин, пролин, серин, треонин, цистеин, метионин, аспартат, аспарагин, глутамат, глутамин, аргинин, гистидин.
Кетогенные: лейцин, лизин.
Глюко-кетогенные (смешанные): изолейцин, фенилаланин, тирозин, триптофан.
«Миллеровские» аминокислоты
Основная статья: Эксперимент Миллера — Юри
«Миллеровские» аминокислоты — обобщенное название аминокислот, получающихся в условиях, близких к эксперименту Стенли Л. Миллера 1953 года. Установлено образование в виде рацемата множества различных аминокислот, в том числе: глицин, аланин, валин, изолейцин, лейцин, пролин, серин, треонин, аспартат, глутамат
12) Свойства аминокислот
Физические свойства. Аминокислоты – твердые кристаллические вещества с высокой т.пл., при плавлении разлагаются. Хорошо растворимы в воде, водные растворы электропроводны. Эти свойства объясняются тем, что молекулы аминокислот существуют в виде внутренних солей, которые образуются за счет переноса протона от карбоксила к аминогруппе.

Химические свойства. Аминокислоты проявляют свойства оснований за счет аминогруппы и свойства кислот за счет карбоксильной группы, т.е. являются амфотерными соединениями. Подобно аминам, они реагируют с кислотами с образованием солей аммония:
H2N–CH2–COOH + HCl Cl [H3N–CH2–COOH]+
Как карбоновые кислоты они образуют функциональные производные:
а) соли
H2N–CH2–COOH + NaOH H2N–CH2–COO Na+ + H2O
б) сложные эфиры
![]()
Кроме того, возможно взаимодействие амино- и карбоксильной групп как внутри одной молекулы (внутримолекулярная реакция), так и принадлежащих разным молекулам (межмолекулярная реакция).Практическое значение имеет внутримолекулярное взаимодействие функциональных групп -аминокапроновой кислоты, в результате которого образуется -капролактам (полупродукт для получения капрона):

Межмолекулярное взаимодействие -аминокислот приводит к образованию пептидов. При взаимодействии двух -аминокислот образуется дипептид.

Межмолекулярное взаимодействие трех -аминокислот приводит к образованию трипептида и т.д.Фрагменты молекул аминокислот, образующие пептидную цепь, называются аминокислотными остатками, а связь CO–NH - пептидной связью.
14) Коллоидные свойства белков Растворы белков обладают свойствами как истиных, так и коллоидных растворов. Это связано с тем, что белки в растворе диспергированы до единичных молекул, но, вследствие большой молекулярной массы и связанного с ней большого размера частиц (1 - 100 нм), растворы белков имеют коллоидный характер.Растворы белка, в связи с коллоидным характером, рассеивают свет (явление Тиндаля), характеризуются высокой вязкостью, при определенных условиях могут терять текучесть и образовывать г е л и, или с т у д н и (студни, сформированные из молекул белков, рассматривают как частную форму гелей).Молекулы белка вследствие большого размера медленно диффундируют в растворе в направлении более низкой концентрации и неспособны проникать через поры искусственных мембран из целлофана, коллодия, пергамента, а также большинства мембран клеток растений и животных. В то же время молекулы низкомолекулярных веществ (вода, этанол, соли, аминокислоты, сахара и т.п.) свободно проходят через такие мембраны.
15)
Амфотерные свойства и изоэлектрическая точка белков
Макромолекулы белков несут на своей поверхности большое количество карбоксильных и аминных групп, что придает им свойства амфотерных полиэлектролитов. Карбоксильные группы, способные к диссоциации с образованием протонов (Н+), определяют кислотные свойства молекулы белка; аминогруппы, способные присоединять протоны, определяют ее основные свойства.Соотношение между количеством кислых и основных группировок варьирует у различных белков. Белки, в которых преобладают кислые группировки, имеют при рН 7 (или близких к 7) суммарный отрицательный заряд и их называют кислыми; белки, в которых преобладают основные группировки, имеют при указанных значениях рН положительный заряд и их называют основными. В живых организмах преобладают кислые белки.Следовательно, изменяя рН среды добавлением кислот или щелочей, можно не только уравнять положительные и отрицательные заряды на поверхности молекулы белка, но и усилить один из них или изменить на противоположный. В кислой среде молекулы белка приобретают положительный заряд и в поле постоянного электрического тока движутся к катоду; в щелочной среде они приобретают отрицательный заряд и в поле постоянного электирческого тока движутся к аноду. Передвижение заряженных растворенных частиц в поле постоянного электрического тока получило название э л е к т р о ф о р е з а (“движение посредством электрического поля”)Для каждого белка (равно пептида и аминокислоты) существует рН при котором положительные и отрицательные заряды в молекуле белка уравновешиваются и суммарный заряд ее становится равным нулю. Такая молекула теряет подвижность в электрическом поле. Величина рН, при котором молекула белка не несет суммарного заряда и не движется в электрическом поле, называется и з о э л е к т р и ч е с к о й т о ч к о й (ИЭТ) и обозначается рНJ; это одна из характерных констант белков. В изоэлектрической точке белок обладает наименьшей растворимостью, легко выпадает в осадок, растворы его менее вязки. Эти явления можно объяснить отсутствием электростатического отталкивания между молекулами белка.
16)
Денатурация белков (от лат. de- — приставка, означающая отделение, удаление и лат. nature — природа; не путать с лат. denaturatus — лишенный природных свойств) — термин биологической химии, означающий потерю белками их естественных свойств (растворимости, гидрофильности и др.) вследствие нарушения пространственной структуры их молекул.
Процесс денатурации отдельной белковой молекулы, приводящий к распаду её «жёсткой» трёхмерной структуры, иногда называют плавлением молекулы. Механизмы денатурацииПрактически любое заметное изменение внешних условий, например, нагревание или обработка белка щелочью приводит к последовательному нарушению четвертичной, третичной и вторичной структур белка. Обычно денатурация вызывается повышением температуры, действием сильных кислот и щелочей, солей тяжелых металлов, некоторых растворителей (спирт), радиации и др.Денатурация часто приводит к тому, что в коллоидном растворе белковых молекул происходит процесс агрегации частиц белка в более крупные. Визуально это выглядит, например, как образование «белка» при жарке яиц. ДЕНАТУРАЦИЯ
ДЕНАТУРАЦИЯ
(от лат. De приставка, означающая удаление, утрату, и nаtura — природные свойства), утрата природной (нативной) конфигурации молекулами белков, нуклеиновых к-т и др. биополимеров в результате нагревания, химич. обработки и т. п. Обусловлена разрывом нековалентных (слабых) связей в молекулах биополимеров (слабыми связями поддерживается пространственная структура биополимеров). Обычно сопровождается потерей оиол. активности — ферментативной, гормональной н др. Может быть полной и частичной, обратимой и необратимой. Д. не нарушает прочных ковалентных химич. связей, но в связи с развёртыванием глобулярной структуры делает доступными для растворителей и химич. реагентов радикалы, находящиеся внутри молекулы. В частности, Д. облегчает действие протеолитич. ферментов, открывая им доступ ко всем частям молекулы белка. Обратный процесс наз. ренатурацией. Обратимую Д. нуклеиновых к-т используют для их молекулярной гибридизации.
Под денатурацией понимают утрату трехмерной конформации, присущей данной белковой молекуле. Это изменение может носить временный или постоянный характер, но и в том, и в другом случае аминокислотная последовательность белка остается неизменной. При денатурации молекула развертывается и теряет способность выполнять свою обычную биологическую функцию. Вызывать денатурацию белков могут разнообразные факторы, перечисленные ниже. Нагревание или излучение белка, например инфракрасное или ультрафиолетовое. Кинетическая энергия, сообщаемая белку, вызывает вибрацию его атомов, вследствие чего слабые водородные и ионные связи разрываются,и белок свертывается (коагулирует). Сильные кислоты, щелочи, соли денатурируют белок. Под действием этих реагентов ионные связи разрываются и белок коагулирует. Длительное воздействие реагента может вызвать разрыв и пептидных связей. Тяжелые металлы денатурируют белок. Положительно заряженные ионы тяжелых металлов (катионы) образуют прочные связи с отрицательно заряженными карбоксил-анионами R-групп белка и часто вызывают разрывы ионных связей. Они также снижают электрическую поляризацию белка, уменьшая его растворимость. Вследствие этого находящийся в растворе белок выпадает в осадок. Органические растворители и детергенты денатурируют белок. Эти реагенты нарушают гидрофобные взаимодействия и образуют связи с гидрофобными (неполярными) группами. В результате разрываются и внутримолекулярные водородные связи. Использование спирта в качестве дезинфицирующего средства основано именно на том, что он вызывает денатурацию белка любых присутствующих бактерий.
Свертывание крови – это важнейший этап работы системы гемостаза, отвечающей за остановку кровотечения при повреждении сосудистой системы организма. Свертыванию крови предшествует стадия первичного сосудисто-тромбоцитарного гемостаза. Этот первичный гемостаз почти целиком обусловлен сужением сосудов и механической закупоркой агрегатами тромбоцитов места повреждения сосудистой стенки. Характерное время для первичного гемостаза у здорового человека составляет 1-3 мин. Свёртывание крови (гемокоагуляция, коагуляция, плазменный гемостаз, вторичный гемостаз) — сложный биологический процесс образования в крови нитей белка фибрина, который полимеризуется и образует тромбы, в результате чего кровь теряет текучесть, приобретая творожистую консистенцию. Свертывание крови у здорового человека происходит локально, в месте образования первичной тромбоцитарной пробки. Характерное время образования фибринового сгустка – около 10 мин.
17) Существует большое количество классификаций белков по различным признакам, но в контексте питания следует рассмотреть его разновидности по степени сложности.
Протеины – простые белки, которые в себе содержат только аминокислотные остатки. Представителями класса являются: альбумины – белок яйца; глобулины – в мышцах, крови, молоке, некоторых видах бобовых и семян; проламины – белки растительных культур (пшеницы, кукурузы, ржи, овса, ячменя); глютелины находятся в семенах пшеницы и риса.
Протеиды – сложные структуры, где в составе имеются и белковые, и небелковые образования. Основные белки этого класса: нуклеопротеиды помимо белковой части располагают нуклеиновыми кислотами; липопротеиды – включают жиры; в фосфопротеидах – присутствует фосфорная кислота.
Существует мнение, протеин важен в первую очередь потому, что он задействован в большом количестве процессов, причем с необыкновенной легкостью и изяществом. Так, наследственная информация, сосредоточенная в белковых молекулах ДНК, характеризует эти природные полимеры как посредника в передаче генетической информации, основы продолжения рода. Без их участия соединения наследственной структуры не способны к процессам воспроизведения и деления. Помимо вышеназванного данные высокомолекулярные вещества выполняют большой спектр функциональных задач.В организме человека белки расщепляются до аминокислот и могут быть заменимыми или незаменимыми[1]. По другой классификации их разнят на виды с природной и синтетической организацией. Первые насчитывают около 150 видов, среди которых 22 разновидности протеиногенных, т.е. содержатся в белках. В отношении высших животных и человека, аминокислоты могут быть:Незаменимые (эссенциальные) аминокислоты не производятся в организме человека автономно, они должны поступать внутрь вместе с животной или растительной пищей. Данное обстоятельство обусловлено тем, что ферментативная система высших животных не позволяет синтезировать радикалы этих соединений. К данному классу белковых остатков причисляют восемь видов: валин, изолейцин, лейцин, лизин[2], метионин, треонин, триптофан и фенилаланин. В таблице представлены суточные нормы каждой из нихКачество белка определяет его биологическую ценность по наличию в нем незаменимых кислот, и является его важнейшим критерием в классификации пищевых продуктов. Чем более изделие насыщено этими компонентами, тем его значимость в организме человека выше.Заменимые аминокислоты образуются в достаточном количестве из поступившего питательного материала. Данный класс составляют 9 видов: аланин, аспаргин, аспаргиновая кислота, глицин, глутамин, глутаминовая кислота, пролин, серин, цистин. Человек способен обходится без них длительный период времени, если только поступают вещества для их формирования.
Некоторые специалисты выделяют еще одну группу полузаменимые аминокислоты, которые вырабатываются организмом, но в недостаточном размере. В связи с этим, рекомендуется дополнительное их применение с пищей. Данную группу составляют: аргинин, гистидин и тирозин. Стоит обратить внимание молодых родителей, что некоторые аминокислоты (например, гистидин) еще не способны синтезироваться в организмах малолетних детей.Несмотря на своё основное предназначение – образование собственных белковых соединений и структур, каждая аминокислота играет специфические роли в значимых жизненных процессах. Рассмотрим некоторые из них:
Глицин участвует в образовании креатина, гемоглобина, серина, пуриновых азотистых оснований, некоторые желчные кислоты, а также обеззараживает токсичные для организма вещества после реакций с участием бензойной и фенилуксусной кислоты.
Серин – основной компонент в производстве веществ, с помощью которых более активно используется гликоген, глюкоза, пировиноградная кислота и цистеин.
Метионин задействован в синтезе витаминных, ферментативных и гормональных соединений: холина, адреналина, креатина, тимина. Более того, без него не обходится и мышечная система.
Триптофан расщепляется либо с формированием ниацина (витамин B5), либо критически важного для жизни серотонина.
Фенилаланин предтеча тирозина, а тот базисное вещество для образования гормонов.
Цистеин – основной фактор образования таурина и цистина, а также некоторых желчных кислот. Аминокислота вовлечена в процесс образования серной кислоты, с помощью которой в печени обезвреживаются ядовитые вещества.
18). ФОСФОПРОТЕИНЫК белкам этого класса относятся казеиноген молока, в котором содержание фосфорной кислоты достигает 1%; вителлин, вителлинин и фосвитин, выделенные из желтка куриного яйца; овальбумин, открытый в белке куриного яйца; ихтулин, содержащийся в икре рыб, и др. Большое количество фосфопротеинов содержится в клетках ЦНС. Фосфопротеины занимают особое положение в биохимии фосфорсодержащих соединений не только в результате своеобразия структурной организации, но и вследствие широкого диапазона функций в метаболизме. Характерной особенностью структуры фосфопротеинов является то, что фосфорная кислота оказывается связанной сложноэфирной связью с белковой молекулой через гидроксильные группы β-оксиаминокислот, главным образом серина и в меньшей степени треонина. На одну молекулу белка обычно приходится 2–4 остатка фосфата.
Рис. 2.4. Ионный тип связи между белками и фосфолипидами.
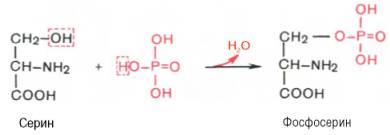
Новые данные свидетельствуют о том, что в клетках фосфопротеины синтезируются в результате посттрансляционной модификации, подвергаясь фосфорилированию при участии протеинкиназ. Этот процесс подробно рассматривается в главе 14. Здесь лишь укажем на существенную роль специфической протеинкиназы, катализирующей фосфорилирование ОН-группы тирозина, в биосинтезе онкобелков. Таким образом, уровень фосфопротеинов в клетке зависит в значительной степени от регулирующего действия ферментов, катализирующих фосфорилирование (протеин-киназы) и дефосфорилирование (протеинфосфатазы). Следует отметить, что фосфопротеины содержат органически связанный, лабильный фосфат, абсолютно необходимый для выполнения клеткой ряда биологических функций. Кроме того, они являются ценным источником энергетического и пластического материала в процессе эмбриогенеза и дальнейшего постна-тального роста и развития организма.Особо следует отметить, что некоторые ключевые ферменты, регулирующие процессы внутриклеточного обмена веществ, также существуют как в фосфорилированной, так и в дефосфорилированной форме. Этим подчеркивается значение фосфорилирования–дефосфорилирования в процессах химической модификации макромолекул, участвующих в интегральных процессах метаболизма.
19)
ЛИПОПРОТЕИНЫ
(липопротеиды), комплексы, состоящие из
белков
(аполипопротеинов;
сокращенно - апо-липопротеины) и липидов,
связь между к-рыми осуществляется
посредством гидрофобных и электростатич.
взаимодействий. Липопротеины подразделяют
на свободные, или р-римые в воде
(липопротеины
плазмы крови,
молока,
желтка яиц и др.), и нерастворимые, т.
наз. структурные (липопротеины мембран
клетки,
миелиновой оболочки нервных волокон,
хлоропластов
растений). Нековалентная связь в
липопротеинах между белками
и липидами
имеет важное биол.
значение. Она обусловливает возможность
своб. обмена липидов
и модуляцию св-в липопротеинов в
организме.
Среди
своб. липопротеинов (они занимают
ключевое положение в транспорте и
метаболизме
липидов)
наиб. изучены липопротеины
плазмы крови,
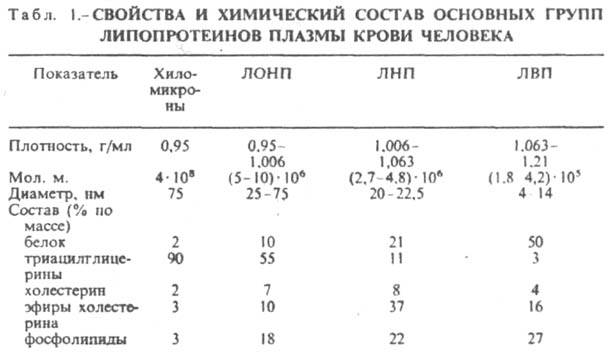
к-рые классифицируют по их плотности.
Чем выше содержание в них липидов,
тем ниже плотность липопротеинов (табл.
1). Различают липопротеины очень низкой
плотности (ЛОНП), низкой плотности (ЛНП),
высокой плотности (ЛВП) и хиломикроны.
Каждая группа липопротеинов очень
неоднородна по размерам частиц (самые
крупные - хиломикроны) и содержанию в
ней апо-липопротеинов. Все группы
липопротеинов плазмы
содержат полярные и неполярные липиды
в разных соотношениях.
 Во
всех липопротеинах плазмы
имеется ядро, состоящее из эфиров
холестерина
и триацилглицеринов, к-рое покрыто
мономолекулярным
слоем
холестерина,
фосфолипидов
и гидрофильной частью молекул
апо-липопротеинов. Предполагают, что
домены
апо-липопротеинов имеют две грани, одна
из к-рых гидрофобная, а другая -
гидрофильная. Первая грань контактирует
с жирно-кислотными цепями фосфолипидов,
вторая - ассоциирована с полярной группой
фосфолипидов
на пов-сти раздела липид
- вода.
Хиломикроны и ЛОНП образуются соотв. в
кишечнике и печени,
ЛВП - в кишечнике, печени,
а также в крови
из др. липопротеинов. Идентифицированы
нек-рые апо-липопротеины (см. табл. 2),
каждый из к-рых обладает определенными
св-вами, связанными с превращением
липопротеинов в организме.
Хиломикроны и ЛОНП, попадая в кровоток,
захватывают апо-липопротеин С-II из ЛВП
и активируют мембраносвязанную
липопротеинлипазу, катализирующую
расщепление в них триацилглицеринов.
Во время катаболизма
ацилглицеринов
апо-липопротеин С-II возвращается к ЛВП,
а хиломикроны и ЛОНП превращаются в ЛНП
- осн. источник клеточного холестерина.
ЛНП поступают во все
клетки
организма,
взаимодействуя со спец. рецептором
мембраны,
активность
к-рого понижается с увеличением
концентрации
внутриклеточного холестерина.
ЛВП, наоборот, способны выводить
холестерин
из клеток.
Предполагают, что высокое содержание
в плазме
ЛНП при низком содержании ЛВП - важный
фактор возникновения атеросклероза
(заболевания, обусловленного образованием
обильных отложений холестерина
и его эфиров на внутр. пов-сти кровеносных
сосудов).
Апо-липопротеин A-I - основной белок
ЛВП. Он активирует сывороточный фермент
лецитин
: холестерол ацилтрансферазу (ЛХАТ),
к-рый катализирует одновременно
расщепление фосфолипидов
и ацилирование
холестерина,
переводя его в биологически неактивную
форму.
Выделяют
липопротеины
плазмы крови
обычно методом флотации,
используя водные р-ры бромидов
Na или К разл. плотности. К
структурным липопротеинам относятся
комплексы липидзависимых ферментов
(напр., аденилатциклаза),
рецспторные комплексы (напр., рецептор
ацетилхолина),
к-рые входят в состав мембран
биологических.
Их строение и биол.
св-ва очень разнообразны
Во
всех липопротеинах плазмы
имеется ядро, состоящее из эфиров
холестерина
и триацилглицеринов, к-рое покрыто
мономолекулярным
слоем
холестерина,
фосфолипидов
и гидрофильной частью молекул
апо-липопротеинов. Предполагают, что
домены
апо-липопротеинов имеют две грани, одна
из к-рых гидрофобная, а другая -
гидрофильная. Первая грань контактирует
с жирно-кислотными цепями фосфолипидов,
вторая - ассоциирована с полярной группой
фосфолипидов
на пов-сти раздела липид
- вода.
Хиломикроны и ЛОНП образуются соотв. в
кишечнике и печени,
ЛВП - в кишечнике, печени,
а также в крови
из др. липопротеинов. Идентифицированы
нек-рые апо-липопротеины (см. табл. 2),
каждый из к-рых обладает определенными
св-вами, связанными с превращением
липопротеинов в организме.
Хиломикроны и ЛОНП, попадая в кровоток,
захватывают апо-липопротеин С-II из ЛВП
и активируют мембраносвязанную
липопротеинлипазу, катализирующую
расщепление в них триацилглицеринов.
Во время катаболизма
ацилглицеринов
апо-липопротеин С-II возвращается к ЛВП,
а хиломикроны и ЛОНП превращаются в ЛНП
- осн. источник клеточного холестерина.
ЛНП поступают во все
клетки
организма,
взаимодействуя со спец. рецептором
мембраны,
активность
к-рого понижается с увеличением
концентрации
внутриклеточного холестерина.
ЛВП, наоборот, способны выводить
холестерин
из клеток.
Предполагают, что высокое содержание
в плазме
ЛНП при низком содержании ЛВП - важный
фактор возникновения атеросклероза
(заболевания, обусловленного образованием
обильных отложений холестерина
и его эфиров на внутр. пов-сти кровеносных
сосудов).
Апо-липопротеин A-I - основной белок
ЛВП. Он активирует сывороточный фермент
лецитин
: холестерол ацилтрансферазу (ЛХАТ),
к-рый катализирует одновременно
расщепление фосфолипидов
и ацилирование
холестерина,
переводя его в биологически неактивную
форму.
Выделяют
липопротеины
плазмы крови
обычно методом флотации,
используя водные р-ры бромидов
Na или К разл. плотности. К
структурным липопротеинам относятся
комплексы липидзависимых ферментов
(напр., аденилатциклаза),
рецспторные комплексы (напр., рецептор
ацетилхолина),
к-рые входят в состав мембран
биологических.
Их строение и биол.
св-ва очень разнообразны
20)
ГЛИКОПРОТЕИНЫ (гликопротеиды),
соед., в молекулах
к-рых остатки олиго- или полисахаридов
ковалентно связаны (О- или N-гликозидными
связями) с полипептидными цепями белка.
Гликопротеины широко распространены
в природе. К ним относятся важные
компоненты сыворотки
крови (иммуноглобулины,
трансферины и др.), групповые в-ва крови,
определяющие групповую принадлежность
крови
человека и животных, антигены
мн. вирусов
(гриппа, кори, энцефалита и др.), нек-рые
гормоны,
лектины,
ферменты.
В образовании О-гликозидной углевод-белковой
связи участвуют гидроксиаминокислоты;
известны комбинации галактоза
(Оа1)-гидроксилизин, галактоза
или арабинозагидроксипролин, но чаще
в узле углевод-белковой связи находятся
серии или треонин.
Известно присоединение остатков
N-ацетилглюкозамина (GlcNAc), маннозы
(Man) и галактозы
к серину,
фукозы
(Fuс) - к треонину,
однако подавляющее большинство таких
гликопротеинов содержит в узле
углевод-белковой связи один и тот же
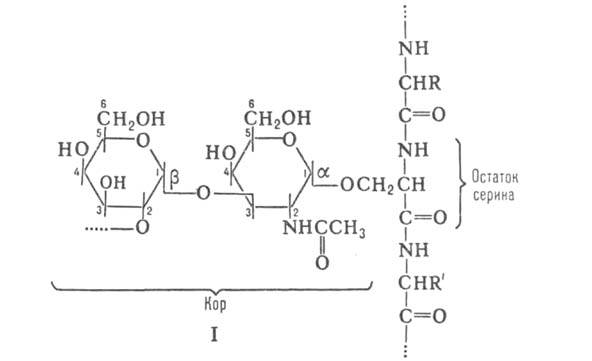
дисахаридный фрагмент (т. наз. кор, или
стержень), к-рый состоит из остатков
галактозы
и N-ацетилгалактозамина (GalNAc) и имеет
строение![]() -(цифры обозначают номера атомов
С в моносахаридах,
участвующих в образовании гликозидных
связей, греч. буквы - конфигурацию
аномерного атома
С). Кор связан с атомом
О гидроксильной
группы остатка серина
или треонина
(см., напр., ф-лу I). В состав углеводных
цепей таких гликопротеинов входят также
остатки N-ацетилглюкозамина, L-фукозы и
(или) N-ацетилнейраминовой к-ты (NeuAc).
-(цифры обозначают номера атомов
С в моносахаридах,
участвующих в образовании гликозидных
связей, греч. буквы - конфигурацию
аномерного атома
С). Кор связан с атомом
О гидроксильной
группы остатка серина
или треонина
(см., напр., ф-лу I). В состав углеводных
цепей таких гликопротеинов входят также
остатки N-ацетилглюкозамина, L-фукозы и
(или) N-ацетилнейраминовой к-ты (NeuAc).
Усложнение
углеводной цепи О-гликозилпротеинов
происходит в результате присоединения
к кору остатков N-ацетиллактозамина![]() ; на конце углеводных цепей находятся
обычно остатки фукозы
или N-ацетилнейраминовой к-ты. Так,
октасахарид, выделенный из групповых
в-в крови,
имеет вид:
; на конце углеводных цепей находятся
обычно остатки фукозы
или N-ацетилнейраминовой к-ты. Так,
октасахарид, выделенный из групповых
в-в крови,
имеет вид:
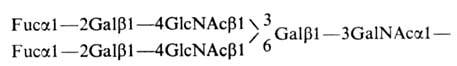
В
более сложном олигосахариде,
выделенном из этого же источника,
дополнит. остаток N-ацетиллактозамина
связан с остатком N-ацетилгалактозамина:
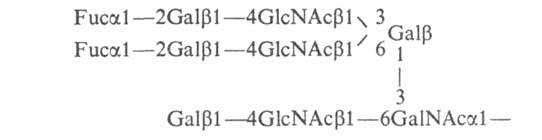
Такие
сложные олигосахаридные цепи обнаружены
гл. обр. в групповых в-вах крови.
Структура углеводных цепей из др.
источников м. б. выражена общей ф-лой:
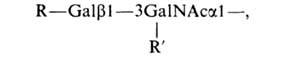
где
R и R'-фукоза и (или) N-ацетилнейраминовая
к-та. Полипептидные цепи О-гликозилпротеинов,
в отличие от обычных белков,
сильно обогащены остатками серина
и треонина.
В гликопротеинах, у к-рых углеводная и
полипептидная цепи соединены N-гликозидными
(гликозил-амидными) связями
(N-гликозилпротеины), остаток
N-ацетилглюкозамина связан с атомом
N амидной группы остатка аспарагина
в полипептидной цепи. В этом случае кор
углеводной цепи гликопротеинов-
пентасахарид:
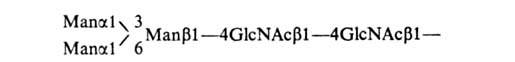
Углеводные
цепи у N-гликозилпротеинов м. б. двух
типов -гликозилманнозные (II) или
комплексные (III):
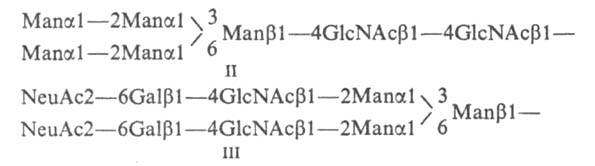
В
состав первых входит только манноза
и N-ацетилглюкозамин. Комплексные цепи
содержат дополнительно галактозу,
а также фукозу
или N-ацетилнейраминовую к-ту и образуются
в результате трансформации
олигоманнозидных цепей при биосинтезе
гликопротеинов. Ветвление углеводных
цепей в О- и N-гликозилпротеинах
осуществляется присоединением к кору
нескольких остатков N-ацетиллактозамина,
в зависимости от числа остатков к-рого
образуются углеводные цепи, содержащие
2, 3 или 4 ветви, или антенны. Напр., в состав
т. наз.![]() кислого
гликопротеина сыворотки
крови входят четырехантенные
углеводные цепи:
кислого
гликопротеина сыворотки
крови входят четырехантенные
углеводные цепи:
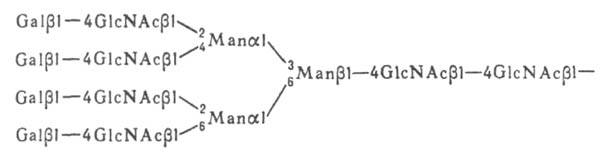
Полипептидная цепь N-гликозилпротеинов по составу аминокислот не отличается от обычных сывороточных белков. Гликопротеины пов-сти клеток ответственны за межклеточные взаимод. и р-ции клетки с внеш. средой. Биол. специфичность О-гликозилпротеинов находится обычно в зависимости от структуры углеводной компоненты молекулы. За биол. специфичность N-гликозилпротеинов (напр., иммуноглобулинов, антигенов вирусов) ответственна в осн. белковая компонента молекулы, а углеводные цепи участвуют гл. обр. в формировании и поддержании необходимой конформа-ции молекулы.
21) Хромопротеиды — строение, представители, значение.
ХП это окрашенные белки (chroma – краска). Молекулы состоят из простого белка и простетической группы, окрашенной обычно за счет металла или витамина. Среди ХП различают дыхательные белки и дыхательные ферменты, которые образуют подгруппу гемопротеидов. Кроме гемопротеидов в группу ХП входят пигменты (родопсин, меланин), магний-порфирины (хлорофилл), желтые ферменты – флавиновые ферменты (выполняют роль дыхательных ферментов).К дыхательным белкам относятся гемоглобин (Нb) – красный пигмент крови и миоглобин (Мgb) – красный пигмент мышц. Хромопротеины – это сложные белки, простетическая часть которых представлена окрашенным компонентом (от греч. Chromos – краска). К ним относятся биологически важные белки гемоглобин, миоглобин, а также некоторые ферменты: каталаза, пероксидаза, цитохромы, все они являются гемпротеинами, так как простетическая часть их содержит гемГемоглобин состоит из простого белка типа гистонов – глобина и 4-х гемов (простетическая группа). Глобин состоит из 2-х п/п альфа цепей (по 141 аминокислоте в каждой) и 2-х бета-цепей (по 146 аминокислот в каждой). Видовая специфичность гемоглобина обусловлена особенностями аминокислотного состава глобина. Например, в глобине человека нет иле. Глобин, соединяясь с гемом, превращает малорастворимую и инертную структуру в хорошо растворимую и активную форму, способную связывать кислород. В свою очередь гемы придают устойчивость большим молекулам глобина.Гем – производное порфирина, состоит из 4-х пиррольных колец, связанных в циклическую структуру метиновыми мостиками. Порфин с заместителями у бета-углерода называется порфирином. Различные порфирины различаются друг от друга характером заместителей. Гемы гемоглобина у 1,3,5,8 атомов углерода содержат СН3 группу (метил), у 2,4 – винильные радикалы, у 6,7 – остатки пропионовых кислот. Соединяясь с ионом железа, порфирин образует гем. железо присоединяется к атомам азота II и IV колец ковалентными и к III и I колец нековалентными (координационными связями). Строение всех 4-х гемов идентичное – показать на табл или пленке и дать списать. Каждый гем соединен с одной п/п цепью (a или b) двумя координационными связями иона железа с имидазольными кольцами гистидинов. Одна из этих связей постоянна, а другая разрывается, когда к гемоглобину присоединяется кислород.
В 1957 году биохимики Д.Кендрью и М.Перутц получили Нобелевскую премию за расшифровку строения молекулы гемоглобина.Во время развития организма гемоглобин претерпевает определенные изменения: на ранних стадиях у эмбриона содержится эмбриональный гемоглобин Е, который после 3-4 месяцев развития заменяется фетальным F, содержащим 2 a и 2 g цепи. Кровь новорожденного содержит 80% фетального гемоглобина, но к концу первого года жизни он почти полностью заменяется на гемоглобин А. В крови взрослого человека все же присутствует 1,5% фетального гемоглобина. Он имеет большее сродство к кислороду, чем гемоглобин взрослого организма – гемоглобин А и обеспечивает снабжение плода кислородом при меньшем его парциальном давлении. В дополнение к основному гемоглобину взрослого человека А1, имеется гемоглобин А2, молекула которого состоит из 2 a цепей и 2 s цепей. На долю гемоглобина А2 приходится 2,5% от всего гемоглобина.
Роль гемоглобина. Гемоглобин – основной белок эритроцитов. В 1-ом эритроците содержится 340 млн молекул гемоглобина, каждая из которых состоит из 103 атомов С, Н, О, N, S и 4 атомов железа. Основная роль – перенос кислорода от легких к тканям (оксигенация – показать на доске) и углекислого газа от тканей к легким. Гемоглобин образует буферные системы, которые участвуют в поддержании КОС. При распаде гемоглобина образуются пигменты кала, мочи и желчи. Гемоглобин участвует в обезвреживании оксида азота, который может присоединяться к нему и образовывать нитрозгемоглобин.Молекула миоглобина состоит из 1-го гема и 1-ой п/п цепи (из 153 аминокислот). Гем миоглобина такой же как у гемоглобина. Роль миоглобина – транспорт кислорода от оксигемоглобина к ферментам дыхательной цепи в клетке – показать на доске реакцию.Содержится, в основном, в цитоплазме мышечных клеток. Также служит в качестве депо кислорода. Миоглобина больше в натренированных мышцах – у диких животных, особенно у ныряющих – кашалота, тюленей (например, у зайца больше, чем у кролика). Миоглобин специфический белок мышц, поэтому его появление вы сыворотке крови говорит о поражении мышечной ткани (заболевания мышц, инфаркт миокарда).
Типы гемоглобинов. Гемоглобины могут различаться по белковой части. Различают физиологические и аномальные типы гемоглобинов. Физиологические типы образуются на разных этапах нормального развития организма, а аномальные - вследствие нарушения последовательности аминокислот в белке глобине физиологических типов гемоглобина. Физиологические типы гемоглобинов отличаются друг от друга набором полипептидных цепей или субъединиц, образующихся на разных этапах развития организма человека – от эмбрионального до взрослого состояния. Различаются следующие физиологические типы гемоглобинов: а) примитивный НвР, появляется на самых ранних стадиях развития эмбриона (1 – 2 недели) Эмбриональный гемоглобин – тетрамер б) фетальный гемоглобин НвF (от лат. Fetus – плод). НвF является главным типом гемоглобина плода и составляет к моменту рождения 70% всего гемоглобина – это тетрамер ( в) гемоглобин взрослого НвА, НвА2, НвА3 ( от лат. Adultus – взрослый ). НвА появляется на более поздних стадиях развития плода, в крови взрослого человека примерно 95 – 96% НвА – это тетрамер (2?, 2?). Гемогловин А2 – тетрамер . Его содержание в эритроцитах взрослого человека равно 2%. Аномальные гемоглобины. Их обнаружено более 200 и отличаются они составом цепей или заменой аминокислот в полипептидных цепях. Из аномальных гемоглобинов часто встречается HвS или серповидно-клеточный Нв. Он обнаруживается у больных серповидно-клеточной анемией. Это широко распространенное заболевание в странах Южной Америки, Африки и Юго-Восточной Азии. При этой патологии эритроциты в условиях низкого парциального давления кислорода принимают форму серпа. Гемоглобин S отличается по ряду свойств от нормального гемоглобина. После отдачи кислорода в тканях он превращается в плохо растворимую форму и начинает выпадать в осадок в виде веретенообразных кристаллоидов. Последние деформируют клетку и приводят к массивному гемолизу.Химический дефект при серповидно-клеточной анемии сводится к замене в белке одной аминокислоты на другую. В норме в b - субъединицах тетрамерной структуры гемоглобина в шестом положении со стороны N – конца находится глутаминовая аминокислота, боковая группа которой имеет отрицательный заряд и характеризуется высокой гидрофильностью. При серповидно-клеточной анемии глутаминовая аминокислота заменена на гидрофобную аминокислоту – валин. Однако одной этой замены оказалось достаточно не только для нарушения формы эритроцита, но и для развития болезни. Миоглобин также относится к хромопротеинам. Это белок, имеющий третичную структуру. Вторичная и третичная структура миоглобина и протомеров гемоглобина очень сходны. Функции миоглобина и гемоглобина одинаковы. Оба белка участвуют в транспорте кислорода. Гемоглобин присоединяет кислород из альвеолярного воздуха и доставляет его в ткани. Миоглобин присоединяет кислород, доставленный гемоглобином и служит промежуточным звеном в транспорте кислорода внутри клетки к митохондриям, а также для запасания кислорода в тканях, создавая кислородный резерв, который расходуется по мере необходимости. В условиях интенсивной мышечной работы, когда парциальное давление кислорода в тканях падает, кислород освобождается из комплекса с миоглобином и используется в митохондриях клеток для получения энергии, необходимой для работы мышц.
22)
Азотистое основание |
Нуклеозид |
Дезоксинуклеозид |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
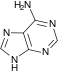 Аденин
Аденин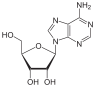 Аденозин
A
Аденозин
A Дезоксиаденозин
dA
Дезоксиаденозин
dA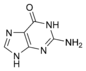 Гуанин
Гуанин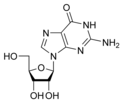 Гуанозин
G
Гуанозин
G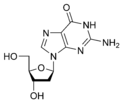 Дезоксигуанозин
dG
Дезоксигуанозин
dG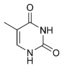 Тимин
Тимин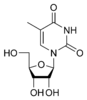 5-Метилуридин
m5U
5-Метилуридин
m5U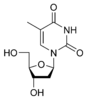 Дезокситимидин
dT
Дезокситимидин
dT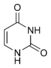 Урацил
Урацил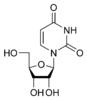 Уридин
U
Уридин
U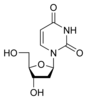 Дезоксиуридин
dU
Дезоксиуридин
dU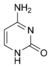 Цитозин
Цитозин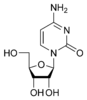 Цитидин
C
Цитидин
C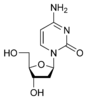 Дезоксицитидин
dC
Дезоксицитидин
dCНуклеозиды могут быть фосфорилированы киназами клетки по первичной спиртовой группе сахара, при этом образуются соответствующие нуклеотиды.[1]
НУКЛЕОЗИДЫ, прир. гликозиды, молекулы к-рых состоят из остатка пуринового или пиримидинового основания, связанного через атом N с остатком D-рибозы или 2-дезок-си-D-рибозы в фуранозной форме; в более широком смысле-прир. и синтетич. соед., в молекулах к-рых гетероцикл через атом N или С связан с любым моносахаридом, иногда сильно модифицированным (см. Минорные нуклеозиды). В зависимости от входящих в молекулу остатков моносахари-да и гетероциклич. основания различают рибо- и дезокси-рибонуклеозиды, пуриновые и пиримидиновые нуклеозиды.
Канонические нуклеозиды (см.рис.)-аденозин (сокращенно А), гуанозин (G), цитидин (С), их 2'-дезоксианалоги, а также тимидин (Т) и уридин (U)-являются компонентами нуклеиновых кислот. В природе нуклеозиды встречаются также в своб. состоянии (преим. в виде нуклеозидных антибиотиков).
Нуклеоти́ды — фосфорные эфиры нуклеозидов, нуклеозидфосфаты. Свободные нуклеотиды, в частности АТФ, цАМФ, АДФ, играют важную роль в энергетических и информационных внутриклеточных процессах, а также являются составляющими частями нуклеиновых кислот и многих коферментов. Нуклеотиды являются сложными эфирами нуклеозидов и фосфорных кислот. Нуклеозиды, в свою очередь, являются N-гликозидами, содержащими гетероциклический фрагмент, связанный через атом азота с C-1 атомом остатка сахара.
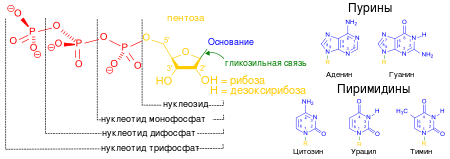
![]()
Строение нуклеотидов
В природе наиболее распространены нуклеотиды, являющиеся β-N-гликозидами пуринов или пиримидинов и пентоз — D-рибозы или D-2-дезоксирибозы. В зависимости от структуры пентозы различают рибонуклеотиды и дезоксирибонуклеотиды, которые являются мономерами молекул сложных биологических полимеров (полинуклеотидов) — соответственно РНК или ДНК.[1]Фосфатный остаток в нуклеотидах обычно образует сложноэфирную связь с 2'-, 3'- или 5'-гидроксильными группами рибонуклеозидов, в случае 2'-дезоксинуклеозидов этерифицируются 3'- или 5'-гидроксильные группы.Большинство нуклеотидов являются моноэфирами ортофосфорной кислоты, однако известны и диэфиры нуклеотидов, в которых этерифицированы два гидроксильных остатка — например, циклические нуклеотиды циклоаденин- и циклогуанин монофосфаты (цАМФ и цГМФ). Наряду с нуклеотидами — эфирами ортофосфорной кислоты (монофосфатами) в природе также распространены и моно- и диэфиры пирофосфорной кислоты (дифосфаты, например, аденозиндифосфат) и моноэфиры триполифосфорной кислоты (трифосфаты, например, аденозинтрифосфат).Соединения, состоящие из двух нуклеотидовых молекул, называются динуклеотидами, из трёх — тринуклеотидами, из небольшого числа — олигонуклеотидами, а из многих — полинуклеотидами, или нуклеиновыми кислотами.Названия нуклеотидов представляют собой аббревиатуры в виде стандартных трёх- или четырёхбуквенных кодов.Если аббревиатура начинается со строчной буквы «д» (англ. d), значит подразумевается дезоксирибонуклеотид; отсутствие буквы «д» означает рибонуклеотид. Если аббревиатура начинается со строчной буквы «ц» (англ. c), значит речь идёт о циклической форме нуклеотида (например, цАМФ).Первая прописная буква аббревиатуры указывает на конкретное азотистое основание или группу возможных нуклеиновых оснований, вторая буква — на количество остатков фосфорной кислоты в структуре (М — моно-, Д — ди-, Т — три-), а третья прописная буква — всегда буква Ф («-фосфат»; англ. P).
Латинские и русские коды для нуклеиновых оснований:
A — А: Аденин;
G — Г: Гуанин;
C — Ц: Цитозин;
T — Т: Тимин (5-метилурацил), не встречается в РНК, занимает место урацила в ДНК;
U — У: Урацил, не встречается в ДНК, занимает место тимина в РНК.
Общепринятые буквенные коды для обозначения нуклеотидных оснований соответствуют номенклатуре, принятой Международным обществом теоретической и прикладной химии (International Union of Pure and Applied Chemistry, сокращённо — англ. IUPAC, рус. ИЮПАК) и Международным обществом биохимии и молекулярной биологии (International Union of Biochemistry and Molecular Biology, сокращённо — англ. IUBMB). Если при секвенировании последовательности ДНК или РНК возникает сомнение в точности определения того или иного нуклеотида, помимо пяти основных (A, C, T, G, U), используют другие буквы латинского алфавита в зависимости от того, какие наиболее вероятные нуклеотиды могут находиться в данной позиции последовательности.Длину секвенированных участков ДНК (гена, сайта, хромосомы) или всего генома указывают в парах нуклеотидов (пн), или парах оснований (англ. base pairs, сокращённо bp), подразумевая под этим элементарную единицу двухцепочечной молекулы нуклеиновой кислоты, сложенную из двух спаренных комплементарных оснований.
3. Выступают в роли коферментов (ФАД, ФМН, НАД+, НАДФ+)
4. Циклические мононуклеотиды являются вторичными посредниками при действии гормонов и других сигналов(цАМФ, цГМФ).
5. Аллостерические регуляторы активности ферментов.
6. Являются мономерами в составе нуклеиновых кислот, связанные 3'-5'- фосфодиэфирными связями
уклеотиды, нуклеозидфосфаты, соединения, из которых состоят нуклеиновые кислоты, многие коферменты и др. биологически активные соединения; каждый Н. построен из азотистого основания (обычно пуринового или пиримидинового), углевода (рибозы или дезоксирибозы) и остатка фосфорной кислоты (см. формулы). Для обозначения дезоксирибонуклеотидов (в отличие от рибонуклеотидов) перед названием Н. обычно ставят букву "д" (например, дАМФ). Соединения, состоящие из остатков двух Н., называются динуклеотидами, из трёх — тринуклеотидами, состоящие из небольшого числа остатков Н. называются олигонуклеотидами, а из многих — полинуклеотидами, или нуклеиновыми кислотами.Н. называются также соединения, построенные по их типу, но содержащие др. азотистые основания (например, никотинамидмононуклеотид — соединение амида никотиновой кислоты с остатками рибозы и фосфорной кислоты). В составе коферментов Н. участвуют во многих процессах обмена веществ [например, никотинамидадениндинуклеотид (НАД) и его фосфорное производное — НАДФ, флавинадениндинуклеотид (ФАД) и др.]. Н. в виде моно-, ди- и трифосфатов содержатся в органах и тканях в свободном состоянии. Н. с тремя остатками фосфорной кислоты — нуклеозидтрифосфаты — являются макроэргическими соединениями и непосредственными предшественниками в биосинтезе нуклеиновых кислот. Особенно важное значение имеет аденозинтрифосфорная кислота (АТФ) — универсальный аккумулятор энергии и непосредственный её источник для различных процессов жизнедеятельности (см. Аденозинфосфорные кислоты). Уридиндифосфорная кислота (УДФ) участвует в углеводном обмене, являясь переносчиком остатков моносахаридов; цитидинфосфаты играют аналогичную роль в обмене липидов; гуанозинтрифосфат (ГТФ) участвует в биосинтезе белка. Важную роль играет также циклическая адениловая кислота (цАМФ), выполняющая роль посредника в действии гормонов.В организме Н. синтезируются из аминокислот и других более простых соединений. Дезоксирибонуклеотиды синтезируются из рибонуклеотидов путём восстановления рибозы. Пуриновые производные у человека окисляются до мочевой кислоты, а у большинства млекопитающих — до аллантоина. Пиримидины распадаются до b-аланина и далее до мочевины.Механизм синтеза пиримидиновых нуклеотидов почти полностью расшифрован благодаря исследованиям П. Рейхарда. Показано, что в клетках животных и в микроорганизмах конечными продуктами синтеза также не являются свободные пиримидиновые основания и остаток рибозы присоединяется к уже сформировавшемуся пиримидиновому кольцу. Синтез начинается с элементарных уровней (СО2, NH3, аспартат), и специфическую ключевую роль выполняет оротовая кислота.
Нуклеозидтрифосфаты (Nucleoside triphosphate, NTP) — это нуклеозиды с тремя фосфатами. Природные нуклеозидтрифосфаты представлены аденозинтрифосфатом (ATP), гуанозинтрифосфатом(GTP), цитидинтрифосфатом(CTP), тимидинтрифосфатом (TTP) и уридинтрифосфатом (UTP). Данные термины означают, что нуклеотиды содержат сахар рибозу.Нуклеозидтрифосфаты (Nucleoside triphosphate, NTP) — это нуклеозиды с тремя фосфатами. Природные нуклеозидтрифосфаты представлены аденозинтрифосфатом (ATP), гуанозинтрифосфатом (GTP), цитидинтрифосфатом (CTP), тимидинтрифосфатом (TTP) и уридинтрифосфатом (UTP). Данные термины означают, что нуклеотиды содержат сахар рибозу.Нуклеотиды, содержащие сахар дезоксирибозу, имеют приставку дезокси- в имени и d- в сокращении: дезоксиаденозинтрифосфатом (dATP), дезоксигуанозинтрифосфатом (dGTP), дезоксицитидинтрифосфатом (dCTP), дезокситимидинтрифосфатом (dTTP) и дезоксиуридинтрифосфатом (dUTP).