

Мембранный потенциал. Движение ионов Cl-
 Мембрана
нейрона проницаема также для ионов Cl-.
Градиент концетрации этих ионов на
мембране противоположен градиенту К+.
Мембрана
нейрона проницаема также для ионов Cl-.
Градиент концетрации этих ионов на
мембране противоположен градиенту К+.
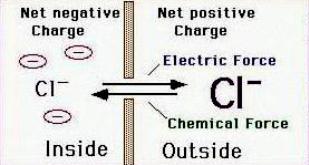
Электрохимическая сила (разница в концентрации иона внутри и вне клетки) приводит к движению Cl- внутрь клетки, отрицательные заряды, попадая в клетку начинают «выталкиваться» электростатической силой, т.к. в клетке много отрицательно заряженных частиц. Потенциал, при котором наступает равновесие этих сил – хлорный равновесный потенциал, приблизительно равный калиевому.
Мембранный потенциал. Движение ионов Na+
 Мембрана
нейрона проницаема также для ионов Na+,
но эта проницаемость в 25 раз ниже.
Градиент концетрации этих ионов на
мембране противоположен градиенту К+.
Мембрана
нейрона проницаема также для ионов Na+,
но эта проницаемость в 25 раз ниже.
Градиент концетрации этих ионов на
мембране противоположен градиенту К+.
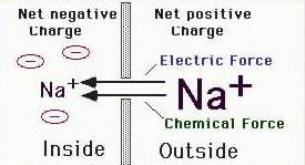
Электрохимическая и электростатическая силы работаютв месте, вызывая движение ионов Na+ внуть клетки. Потенциал равновесия Na+ имеет противоположный знак по отношению к
равновесному потенциалу К+ и, несмотря на низкую проницаемость мембраны для натрия, эти потенциалы должны были бы уравновесить друг друга. Этого не происходит, благодаря существованию специального активного механизма, осуществляющего движение ионов Na+ и K+ против их градиентов концентрации.
Равновесные диффузионные потенциалы. Уравнение Нернста.
 A
(эл.х)=2,3
RT lg([K+]out / [K+]in)
A
(эл.х)=2,3
RT lg([K+]out / [K+]in)
A (эл.)= FE
A (эл.х) = A (эл.)
FE = 2,3 RT lg([K+]out / [K+]in)
E k = -75 мВ
Равновесный потенциал для данного иона определяется соотношением его вне- и внутриклеточной концентрации, а также газовой постоянной (R) (мерой внутренней энергии вещества), температурой (T), постоянной Фарадея (F) (количеством электрических зарядов в 1 моле вещества)
ПП – это отрицательный заряд цито-плазмы, останавли-вающий диффузию ионов К+ в межклеточную среду.
Как долго идет диффузия К+ из нейрона?
Очевидный вариант («до выравнивания концентраций») неверен, поскольку двигаются заряженные частицы, и выход К+ сопровождается накоплением в цитоплазме отрицательного заряда.
«Уравнение
Нернста»: ПП ~ lg
( К+out
/
К+in
) (пропорционально)
)
коэффициент пропорциональности равен 61.5 мВ для Т=36.6°С
логарифм равен -1.48 (для соотношения концентраций 1/30). С учетом этого ПП = -91 мВ («равновесный потенциал» для К+)
В реальной клетке ПП находится ближе к нулю (в среднем -70 мВ). Причина: существование небольшого количества постоянно открытых каналов для ионов Na+.
Избыток ионов Na+ в межклеточной среде, а также их притяжение к отрицательно заряженной цитоплазме приводят к входу Na+ в клетку.
Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверх и частичной потере ПП (отсюда название – «ток утечки Na+ »).
Ограничивает вход Na+,
во-первых, малое число постоян-но открытых Na+-кана- лов;
во-вторых, работа Na+-K+- АТФазы, которая «откачивает» Na+, обменивая его на К+.
В целом ПП зависит от 3-х главных факторов:
- диффузии K+ из клетки;
- диффузии Na+ в клетку;
- работы Na+-K+-АТФазы.
Исходное состояние нейрона (до развития ПП) — отсутствие внутреннего заряда. Количество положительных и отрицательных ионов в цитоплазме клетки равное. Такая картина наблюдается на ранних этапах эмбрионального развития нервной ткани. Затем по мере ее созревания включаются гены, запускающие синтез постоянно открытых К+-каналов. После их встраивания в мембрану ионы К+ получают возможность свободно выходить из клетки, где их содержание велико, в межклеточную среду, где их гораздо меньше.
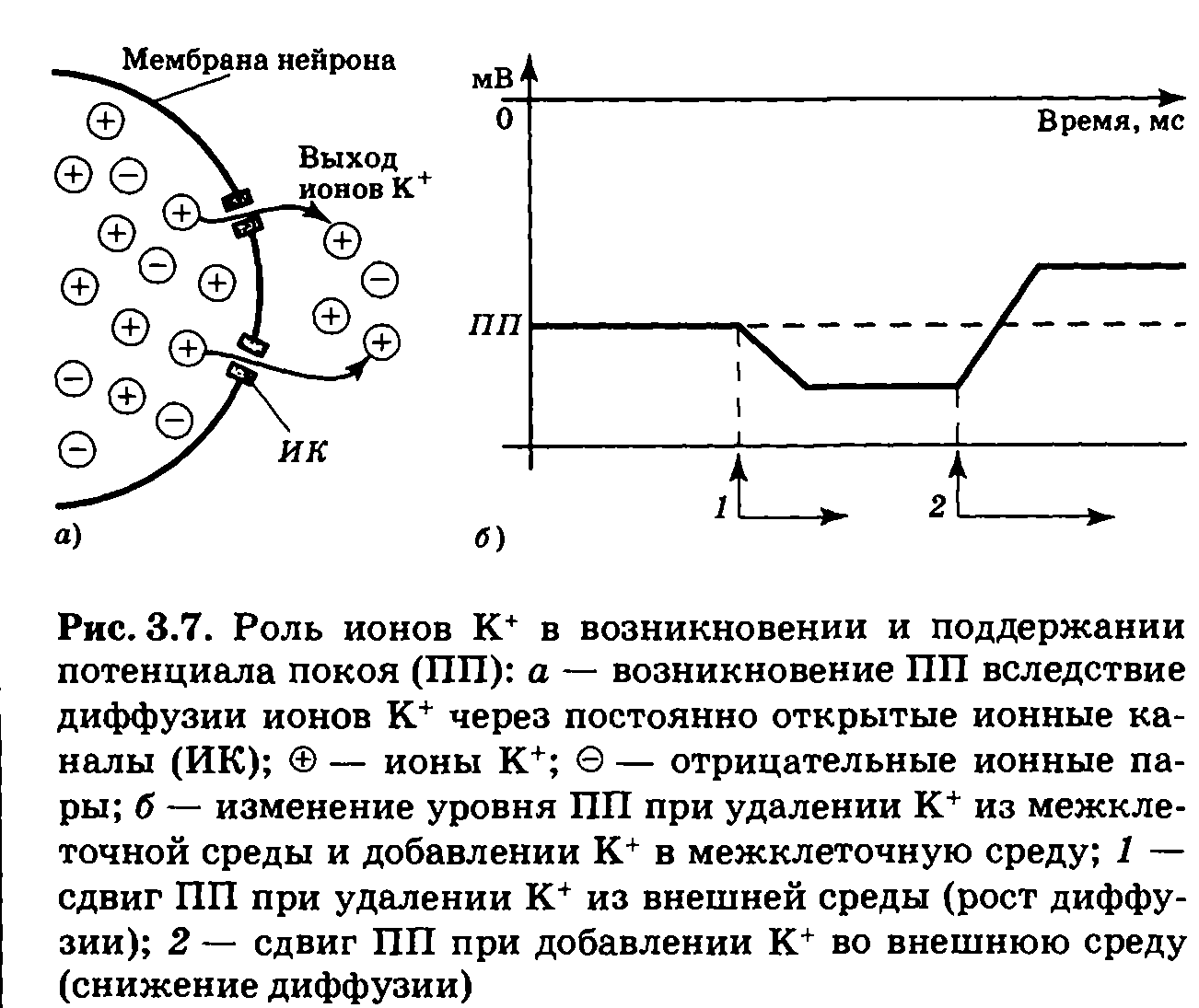
Однако такая диффузия не приводит к уравниванию концентрации ионов калия внутри и вне клетки, так как параллельно с выходом К' из клетки в ней становится все больше оставшихся без своей положительно заряженной пары анионов; по мере диффузии К+ в цитоплазме нейрона накапливается отрицательный электрический заряд (рис. 3.7, а). Притягивая к себе катионы, он начинает все больше препятствовать выходу К+ из клетки. В итоге выход К+ продолжается лишь до тех пор, пока сила диффузии (концентрационного давления) и сила электрического поля не станут равны. Точка равновесия и соответствует уровню потенциала покоя.
Ток ионов при достижении ПП не останавливается, так как имеются постоянно открытые каналы и K+ продолжает перемещаться сквозь мембрану, но число ионов, которые вошли в клетку и вышли из нее, теперь оказывается одинаковым. Такое состояние называется динамическим равновесием — равенством двух противоборствующих процессов. Если интенсивность одного из процессов изменится, то точка равновесия сместится: при искусственном повышении концентрации К+ в межклеточной среде ослабляется диффузия; часть ионов будет втянута внутрь клетки, и ПП сместится вверх. При уменьшении содержания ионов калия в межклеточной среде усиливается диффузия, и ПП сместится вниз (рис. 3.7,б). Все эти явления подчиняются химическим и физическим законам.
Существует уравнение Нернста, позволяющее рассчитать уровень ПП, зная внутреннюю и наружную концентрации К+:

где R, Т и F — три физические характеристики (газовая постоянная, абсолютная температура и постоянная Фарадея соответственно); выражение перед логарифмом 2,3 RT/F) при температуре тела равно 61,5 мВ. Десятичный логарифм отношения концентраций ионов К+ в межклеточной среде ([К+]нар) и цитоплазме ([К+]внутр) равен -1,30 для соотношения 1/20 и -1,48 для соотношения 1/30; соответствующие значения ПП будут равны -80 мВ и -91 мВ; при этом в равновесном состоянии из
клетки выйдет менее 1% ионов К+.
Однако возникает несоответствие между рассчитанным уровнем ПП и ПП реальных клеток, который обычно несколько ниже по абсолютному значению. Подъем ПП происходит за счет другого типа постоянно открытых ионных каналов — натриевых, которых существенно меньше, чем калиевых, но входящие в них ионы Na+, внося в клетку положительный заряд, могут увеличить ПП.
Входящий ток Na+ (ток утечки), снижая ПП по абсолютному значению, делает нейрон более возбудимым. Чем больше у нейрона ток утечки Na+ (постоянно открытых Na+-каналов), тем легче вызвать у него потенциал действия. Роль ионов К+ в формировании ПП состоит в создании «базовой», несколько завышенной разности потенциалов между цитоплазмой нейрона и внешней средой, роль ионов Na+— в «подгонке» ПП каждого конкретного нейрона к генетически заданному уровню возбудимости. Чем больше натрия во внешней среде, а в мембране — натриевых каналов, тем выше (меньше по абсолютному значению) уровень ПП. Таким образом, возникает возможность сформировать как легко возбудимые нейроны (необходимые для систем бодрствования или кратковременной памяти), так и нейроны мало возбудимые (например, запускающие двигательные программы).
Ток утечки Na+ компенсируется работой Na+, К+-насоса. При отравлении строфантином уровень ПП начнет медленно смещаться вверх, приближаясь к нулю.
Мембрана нервной клетки на уровне ПП обладает также определенной проницаемостью для ионов Сl-. Обычно она несколько выше, чем проницаемость для ионов Na+, и вклад ее в формирование ПП учтен в расширенном варианте уравнения Нернста:

где Рк, PNa и РС1— проницаемости мембраны для соответствующих ионов (примерное соотношение 1,00 : 0,04 : 0,15).
Последний фактор, который влияет на уровень ПП, — это электрогенный эффект Na+, К+-насоса. Обменивая в каждом цикле три иона натрия на два иона калия, насос уменьшает общее количество положительно заряженных ионов в цитоплазме, и его активность снижает уровень ПП (увеличивает его по абсолютному значению). Вклад электрогенного эффекта насоса зависит от тока утечки Na+, но обычно не превышает нескольких мВ.
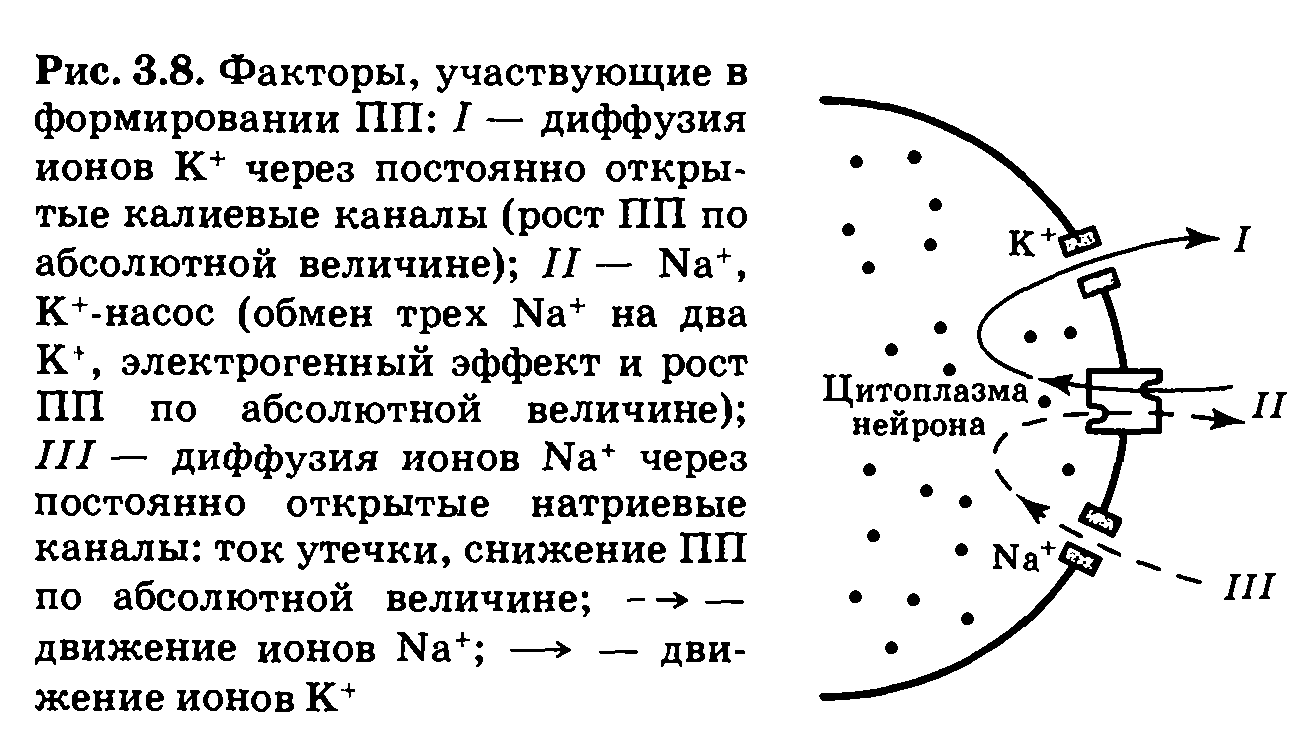
Таким образом, конечный уровень потенциала покоя определяется взаимодействием большого числа факторов, основными из которых являются входящий и выходящий токи К+, входящий ток Na+ и деятельность Na+, К+-насоса. Конечная величина ПП является результатом динамического равновесия процессов. Воздействуя на любой из них, можно смещать уровень ПП и соответственно уровень возбудимости нервной клетки (рис. 3.8).
Таким образом, функция ПП состоит в стабильном обеспечении работоспособного состояния нервных клеток, и, обладая ПП, нейрон способен из состояния ожидания переходить в «рабочий режим» — т. е. время от времени генерировать потенциалы действия (скачки потенциала на мембране, передающие информацию в ЦНС).
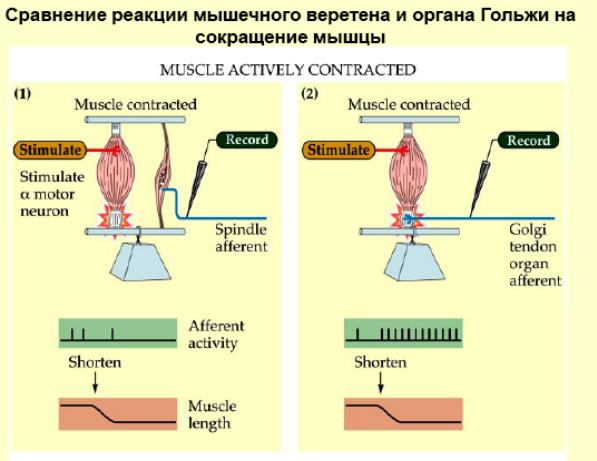
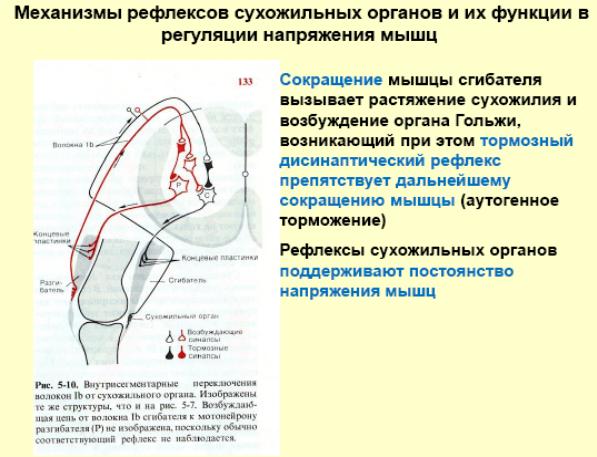
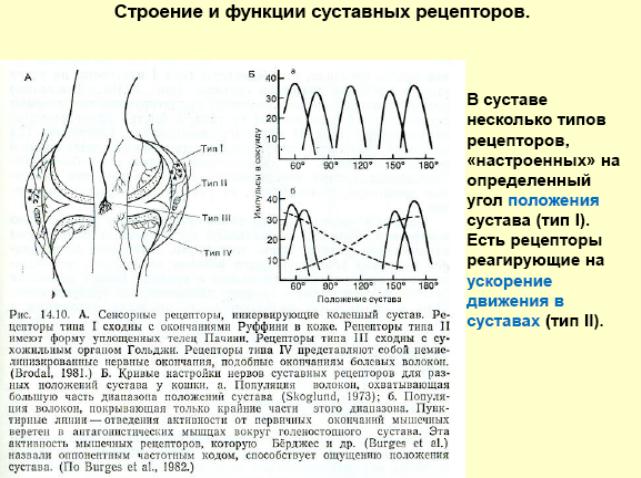
Строение и функции мышечного веретена и сухожильного органа Гольжи. Сравнение реакции мышечных веретен и сухожильных органов на сокращение мышцы. Организация рефлексов растяжения и их роль в обеспечении позы. Механизмы сухожильных рефлексов и их роль в регуляции мышечного напряжения.
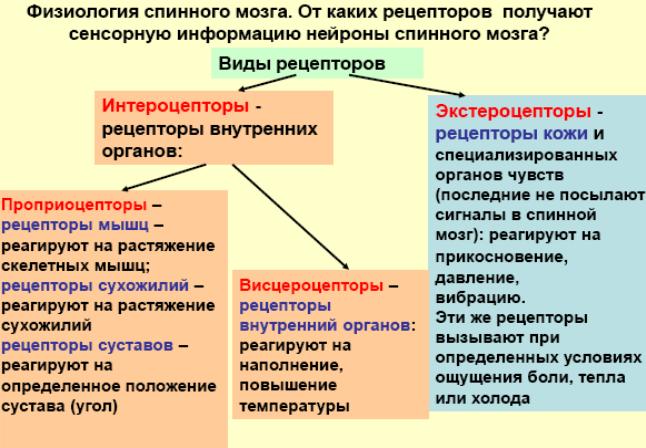
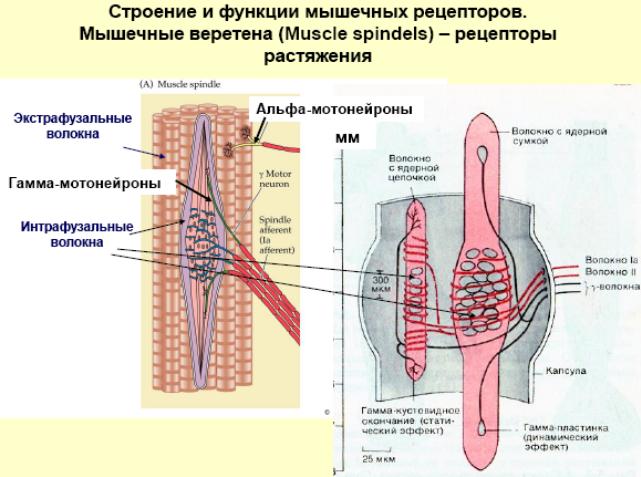
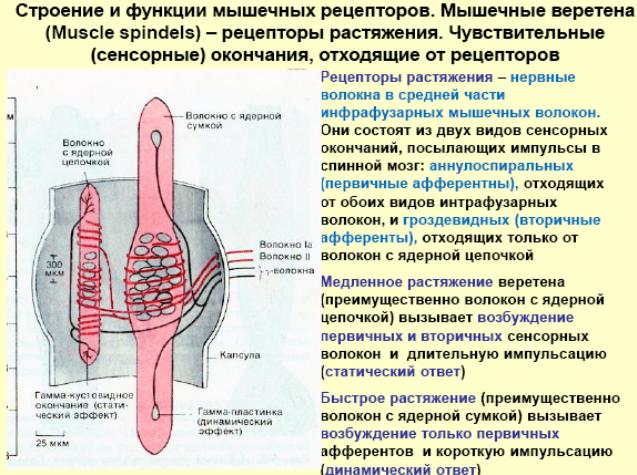

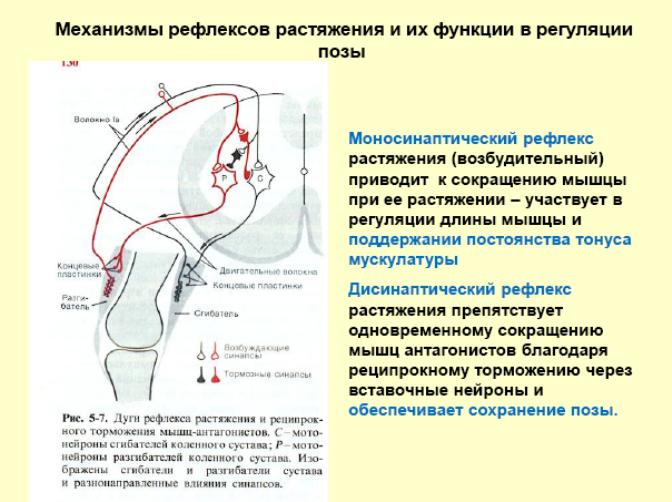

Сухожильный орган Гольджи (нервно-сухожильное веретено) — рецепторный орган, располагающийся в местах соединения мышечных волокон с коллагеновыми пучками сухожилий.
Весь орган окружён тонкой соединительнотканной капсулой. Нервное волокно, подходящее к веретену утрачивает миелиновый слой и, распадаясь на терминальные ветви, проходит между пучками коллагеновых волокон сухожилия.
Сокращаясь, мышца растягивает сухожилие. Это вызывает возбуждение нервных окончаний чувствительных нейронов спинного мозга. Они возбуждают тормозные нейроны спинного мозга, которые тормозят соответствующие двигательные нейроны, что предотвращает перерастяжения мышцы. Известно, что порог возбуждения рецепторов сухожильного органа Гольджи при механическом растяжении мышцы выше, чем аналогичный порог возбуждения мышечных веретен. Это позволяет мышце сокращаться без всяких помех до определенного предела.
Нервно-мышечное веретено — сложный рецептор, который включает видоизмененные мышечные клетки, афферентные и эфферентные нервные отростки и контролирует как скорость, так и степень сокращения и растяжение скелетных мышц.
Достаточно нанести легкий удар по сухожилию мышцы (лучше разгибательной), как последняя отвечает коротким и быстрым сокращением. Общеизвестным примером сухожильного рефлекса является коленный рефлекс, возникающий при ударе по сухожилию четырехглавой мышцы бедра; аналогичные рефлексы можно получить при ударе по ахиллову сухожилию (в этом случае сокращается икроножная мышца) и по сухожилию мышц передних конечностей.
Если таким образом и сухожильные рефлексы и рефлексы растяжения связаны с возбуждением рецепторов мышечных веретен, то какова же функция рецепторов, расположенных в сухожилиях и, в частности, рецепторов Гольджи? Есть основания считать, что возбуждение рецепторов Гольджи является причиной аутогенного торможения собственных рефлексов мышц. Возбуждение афферентных волокон, которые идут от рецепторов Гольджи, вызывает развитие тормозящих постсинаптических потенциалов в соответствующих мотонейронах и прекращает рефлекторный тонус мышцы при ее чрезмерном растяжении. Известно, что порог возбуждения рецепторов Гольджи при механическом растяжении мышцы выше, чем аналогичный порог возбуждения мышечных веретен. Это различие хорошо согласуется с теми условиями раздражения, при которых развивается аутогенное торможение.