

Меры борьбы
Предупреждение вшивости среди населения и организованных коллективов, истребление вшей в случае их появления составляют основу профилактики сыпного и возвратного тифов, а также волынской лихорадки. Каждый случай завшивленности должен рассматриваться как чрезвычайное происшествие, ведущее к проведению комплекса специальных мероприятий. Одним из важных мероприятий по предупреждению вшивости является регулярное санитарно-эпидемиологическое обследование населения. В организованных коллективах (детские сады, ясли, воинские подразделения и т.п.) осмотру на вшивость с целью раннего выявления завшивленности подлежат волосистые части тела, внутренняя поверхность воротников одежды, белья, швы белья под мышками, пояс белья и брюк и др. Мытье тела, смене нательного и постельного белья не реже I раза в 7—10 дней, регуляр-
1шл СТрпЖКа, ОПрлТКОС СОДСрЖаНИС ОДСЖДЫ И "у'хгОрКа ПймСЩСпйм ЯВЛЯЮТСЯ благоприятными факторами в профилактике педикулеза. Важное значение имеет саннтарно-просветительная работа среди населения.
По эпидемическим показателям с целью профилактики паразитарных тифов в организованных коллективах проводят импрегнацию (пропитку) нательного белья водной эмульсией карбофоса. Белье из бязи или полотна замачивают в течение 5 мин в 0,15%-ной водной эмульсии карбофоса; пропитку белья из фланели и трикотажа осущес-
256
твляют путем 3-минутного замачивания в 0,1%-ной водной эмульсии карбофоса, после чего белье хорошо отжимают и высушивают. Норма расхода эмульсии на комплект нательного белья составляет 800—100 мл. Для удаления запаха импрегнированное белье проветривают в течение 6 сут на открытом воздухе или в хорошо вентилируемом нежилом помещении, затем его проглаживают утюгом. Инсектицидные свойства белья в процессе носки сохраняются около 15 дней.
При появлении вшивости среди коллектива проводится одномоментная санитарная обработка, предусматривающая помывку всех людей в санитарном пропускнике с обработкой волос и сменой нательного белья, дезинфекция белья и одежды, уничтожение вшей в помещениях.
При педикулезе применяют инсектициды, обладающие овицид-ным действием: ацетофос, метил-ацетофос, 3-5% которых добавляют к жидкому мылу, шампуню, одеколону или вазелину. При головном и платяном педикулезе в очагах заболеваний или в тех случаях, когда отсутствуют другие средства, допускают использование бутадиона — синтетического лечебного препарата. При приеме его внутрь кровь человека становится токсичной для вшей примерно 14 сут. Головные вши более чувствительны к бутадиону. Этот препарат назначают взрослым после еды в течение 2 дней по 0,15 г 4 раза в день. Суточная доза не должна превышать 0,6 г. Детям в возрасте до 4 лет применение бутадиона противопоказано; от 4 до 7 лет его назначают по 0,05 г; от 8до 10 лет — 0,08 г; старше Шлет — 0,12 г; указанные дозы рекомендуют принимать 3 раза в течение 2 дней.
Ш. МЕДИЦИНСКАЯ АКАРОЛОГИЯ Общая характеристика клещей
Клещи представляют собой особую группу членистоногих, которые относятся к подтипу хеяицеровых (Chelicerata), классу паукообразных (Arachnida). Они различны по морфологии, жизненному циклу географическому распространению, медицинсконги "уивч^иию Наибольшее значение клеши имеют как переносчики и хранители в природе возбудителей инфекционных болезней человека и животных. Главными из них служат представители двух отрядов: Parasitiformes и Acariformes (паразитиформные и акариформные клещи) (таб.13). Отряд Parasitiformes включает два надсемейства — Ixodoidea и Gamasoi-dea. Первое состоит из семейств Ixodidae и Argasidae, второе — из многих семейств.
Иксодоидные клещи (Ixodoidea) являются облигатными крово-
17-1270 257
Таблица J 3
Систематика клещей — переносчиков возбудителей болезней человека
Тип Членистоногие — Arthopoda Подтип Хелицсровыс — Chelicerata Класс Паукообразные — Arachnida
|
|
|
Ixodes Haemaphysalis |
|
|
Икоодовые Ixosidae |
Hyahmma Dermacentor |
|
Иксодоидные Ixodoidea |
|
Rhipkephalus BoophUus |
|
|
Ormthodoros |
|
|
|
Аргасовые Argasidae |
Alveonasus Argas Otobius |
Паразитформные |
|
Haemogamasidae |
Haemogamasus |
Parasitiformes |
|
Eulaelaps |
|
|
|
|
Haemolaelaps |
|
|
Laeiaptidae |
Laelaps Hyperiaelaps |
|
Гамаэондные Gamasoidea |
|
Myonyssus |
|
Hirstionysstdae |
Hirstionyssus |
|
|
Macronyssidae |
Ornithonyssus Pellonyssus |
|
|
Dermanyssidae |
Dermanyssus Allodermanyssus |
|
Акариформные Acariformes |
Тромбидиформ- ные ТготЬеа |
Краснотелкн Trombiculidae |
Leptotrombidium Neotrombicula |
сосами, паразитирующими, эа небольшим исключением, во всех фазах развития (у подавляющего большинства — оба пола) на наземных позвоночных животных. На хозяев они нападают периодически, постоянных паразитов среди них нет. Питаются данные клеши только кровью и лимфой.
Все нксодоидные клеши во взрослом голодном состоянии имеют крупные размеры (1—12мм и болеев длину). Тело их разделено на два отдела: лишенное сегментации туловище (или идиосому), несущее ноги, и гнатосому, или хоботок. Идиосома обычно овальная, у голодных особей более или менее уплощеная, у сытых — значительно раздувшаяся. По краю тела могут располагаться крупные фестоны или рант из мелких складок покровов. Цвет тела у клещей может быть от песочного до красно-коричневого или даже черного, иногда со светлым рисунком.
Покровы клещей образованы однослойной гиподермой, покрытой снаружи кутикулой из нескольких слоев разной структуры н химического состава. Они могут быть мелкоскладчатыми, бугорчатыми и т.п. За счет расправления складок и других образований покровы могут значительно растягиваться. У форм с удлиненным сроком кровососания, всасывающих большие порции крови в процессе питания, покровы способны расти и утолщаться. Отдельные участки кутикулы могут иметь особую структуру и в виде небольших плоских «дисков» располагаться определенным образом на спинной н брюшной сторонах тела клеща (аргасовые клещи). Это места прикрепления мышц к внутренней стороне покровов. В других случаях участки кутикулы сильно склеротнрованы и образуют крупные щитки (спинной и брюшные). На брюшной и спинной сторонах находятся разного рода бороздки и складки, форма и расположение которых имеют значение в систематике клещей.
Ноги клещей состоят из 6 члеников: тазик, вертлуг, бедро, голень, предлапка и лапка. Первая пара конечностей представлена хе-лицерами, служащими для схватывания, жевания или прокалывания пищи, вторая пара — педипальпами; остальные пары конечностей — ходильные ноги.
В отличие от насекомых у клещей отсутствуют антенны. Хобо-то* у нкглдпидт.п[ кттептей приспособлен не только для приема пиши. но и для присасывания и удержания на хозяине в период кровососания. Он состоит из основания, непарного гипостома (длинного, массивного выроста основания гнатосомы, который покрыт обратно направленными зубчиками), парных хелицер и щупиков педипальп (пальп). Хелицеры 2-члениковые, несут по наружному краю мощные режущие зубцы и в покое втянуты внутрь трубчатых футляров; паль* пы — 2-члениковые, прикрывают гипостомы и футляры хелицер сверху и с боков. Формы частей хоботка варьируются у разных предста-
вителей семейств.
Пищеварительный аппарат у иксоидных клещей представлен предротовой полостью, парой слюнных желез, глоткой, гшщеводом, средней и задней кишками (рис.82). Средняя кишка включает небольшой желудок и обширные боковые выросты, задняя кишка — короткую тонкую кишку, ректальный пузырь и прямую кишку, открывающуюся наружу анальньш отверстием. В ректальный пузырь попадает пара мальпигиевых сосудов. Пищеварение у клещей в основном внутриклеточное, происходит главным образом в средней кишке. У иксо-довых клещей клетки этой кишки захватъшают всю поглощенную
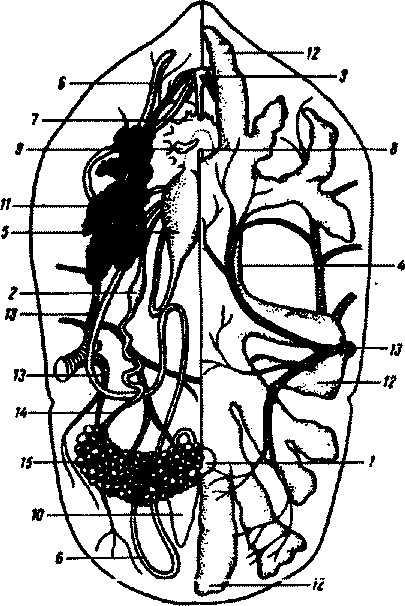
Рис.82. Внутреннее строение самки клеща Ormthodorspapillipes. Левая полови на кишечника удалена (по Ю.С.Балашову):
/ - анус; 2 - ампульное расширение яйцевода; 3 - глотка; 4 - желудок; 5 матка; б - мальпигиев сосуд; 7 - мозг; 8 - пищевод; 9 - придаточная железа; 10 - ректальный пузырь; 11 - слюнная железа; 12 - отростки средней кишки; 13 - трахейные стволы; 14 - яйцевод; 15 - яичник
260
пищу, у аргасовых — лишь большую ее часть, а остальная остается в просвете кишечника и постепенно переваривается секретом, поступающим из разрушающихся секреторных клеток.
Питание начинается с внедрения хоботка в кожные покровы хозяина. Процесс этот протекает сравнительно медленно, путем последовательного прорезания покровов хелицерами и введения в ранку гипостома. Пальпы остаются на поверхности покровов и в акте сосания прямого участия не принимают. Насасывание крови и лимфы чередуется с введением в ранку слюны. Слюна обладает свойствами обезболивать укус, увеличивать проницаемость стенок кровеносных сосудов, предотвращать свертывание крови, может обеспечивать растворение тканей, участвовать в образовании цементного футляра вокруг хоботка.
Органами выделения у клещей, как и у насекомых, служат маль-пигиевы сосуды, через которые удаляются продукты азотистого обмена и осуществляется осморегуляторная функция. Выделение избытка воды у иксодовых и аргасовых клещей происходит за счет «откачивания» его со слюной в ранку, благодаря чему кровь в кишечнике концентрируется. Однако необходимо иметь в виду, что процесс этот многократно повторяется: выделение слюны, ее обратное всасывание вместе с кровью и т.д. Аргазиды, кроме того, избыток воды при кро-вососании выделяют через коксальные железы.
Кровеносная система у иксодоидных клещей представлена сердцем в виде небольшого треугольного мешка с 1—2 парами отверстий, снабженных клапанами, короткой аортой, открывающейся в кровеносный синус у мозга, окологлоточным синусом и несколькими парами кровеносных сосудов, идущих от синусов к конечностям и гнатосо-ме. Гемолимфа бесцветна, содержит амебовидные гемоциты.
У нимф и имаго иксодоидных клещей хорошо развита трахейная система, охватывающаяся парными стигмами на перитремах. Стигмы снабжены замыкательным аппаратом. У личинок большинства видов трахейной системы нет.
Половой аппарат у самок клещей состоит из непарного яичника, парных яйцеводов, сливающихся в непарную матку, и влагалища. Во влагалище открываются семяприемник и пара придаточных половых желез. Иногда семмлрисмши. отсутствует и его функции выполняют матка или влагалище. Половой аппарат самцов включает пару семенников и семяпроводов, семяизвергательный канал и комплекс придаточных желез. Копулятивные органы отсутствуют. Оплодотворение происходит при помощи сперматофора. Самцы вводят шейку сперма-тофора в половые пути самок с помощью хелицер.
Развитие яйцеклеток стимулируется приемом крови. Созревают они в разных участках яичника не одновременно, поэтому яйцекладка у клещей растянута. В процессе яйцекладки важную роль играет орган
26!
Жене, расположенный в передней части тела. Он представляет собой 2- или 4-лопастный пшодермальный мешок с железистой тканью. В момент яйцекладки он выворачивается наружу через зделевидное отверстие у заднего края основания хоботка, захватывает и переносит яйца от полового отверстия к передней части тела клеща и на субстрат. Яйца откладываются в общую кучку впереди самки. Железы органа Жене выделяют секрет, которым покрываются откладываемые яйца.
Жизненный цикл клещей включает фазы яйца, личинки, нимф и имаго. Яйца округлой или овальной формы, желтовато-коричневые, блестящие. Личинки мелкие, с 3 парами ног, без перитрем и полового отверстия. Спинной щиток клещей имеет различную форму. Нимфы крупнее личинок, имеют 4 пары ног, развитые перитрсмы, но еще без полового отверстия. Нимф в жизненном цикле может быть одна или несколько (у разных групп клещей). Имаго отличается от нимф большими размерами, наличием полового отверстия.
Размеры гамазоидных клещей (Camasoidea) значительно меньше, чем иксодоидных,—от 0,7 до 2,6 мм (редко 5 мм). Форма тела чаще овальная. На спинной поверхности взрослых клещей находится ряд щитков, имеющих разную форму. Кутикула личинок и нимф чаще бесцветная, почти прозрачная, щитки на теле отсутствуют или неясно намечены. На теле клещей имеется ряд щетинок, расположение и форма которых разнообразны.
Клешни хелицер у гамазид отличаются от таковых прочих ме-зостигматичесхих клещей: у них подвижный палец часто несет особый вырост — сперматодактиль, отсутствующий у других клещей. Генн-тальное отверстие самца расположено близко к передней части идиосомы. Стигмы дейтонимф и взрослых клещей имеют узкие длинные перитремы; у протонимф перетремы короткие.Среди всех паразита-формных клещей гамазовые клещи наиболее многочисленная группа по количеству видов и наиболее разнообразная в экологическом отношении. Среди них много как свободноживущих (преимущественно хищных или многоядных), так и паразитических видов. Гамазовым клещам свойственны 2 способа осеменения: токоспермия (половые элементы самца вводятся в вагинальную полость) и подоспермия (осеменение при помощи сперматодактиля через парные отверстия — гоно-поры). Строение спёрмахек ичснь рюмиибр&зно. V самки одновременно созревает I яйцо. Клещи могут быть как яйцекладущими, так и живородящими. Метаморфоз бинимфальный; обычно нимфы I и II заметно отличаются друг от друга (лишь у некоторых паразитических клещей нимфы почти не различимы). Малая плодовитость гамазид компенсируется быстротой развития на всех фазах.
Отряд Acariforates включает два подотряда: саркоптнформные клещи (Sarcoptifonnes) и тромбидиформные клещи (Trombidiformes), морфология которых наиболее примитивная. Эти клещи очень малы,
262
всего 0,2—0,3 мм длиной. Туловище их редко сегментировано. Важную роль играют вояоски, щетинки и другие кожные органы, поскольку они выполняют функции осязательных (трихоботрии на переднем отделе тела) и обонятельных органов чувств. Обонятельные представлены лировидными органами. Хелицеры и педипальпы свободные, не соединяются в «головку», обычно 2-члениковые, с клешней. Тазики . педипальп подвижные и имеют жевательные лопасти. Ноги у саркоп-тиформных клещей сильно изменены.
Акариформные клеши лишены специальных органов дыхания и дышат кожей. Оплодотворение у них сперматоформное. Самец оставляет на субстарате клейкие сперматофоры на ножке, а самка захватывает их наружными половыми органами.
Клещи имеют важное медицинское значение. Одни из них (например, чесоточный клещ) вызывают заболевания в силу паразитического образа жизни, другие служат переносчиками возбудителей болезней человека. Роль клещей как переносчиков возбудителей трансмиссивных болезней усиливается в связи с тем, что они одновременно являются природными резервуарами возбудителя. В силу большой индивидуальной долговечности, полифагии и исключительной способности к трансовариальной и трансфазовой передаче микроорганизмов клещи обеспечивают устойчивое поддержание природных очагов трансмиссивных болезней как при непрерывной циркуляции возбудителя между клещами и позвоночными, так и при длительном отсутствии восприимчивых позвоночных животных.
Типы взаимоотношений клещей с патогенными для позвоночных микроорганизмами отличаются исключительным разнообразием. В ряде случае (при заражении туляремийными микробами или филяри-ями) микробы приводят к гибели клеща, а в случае с риккетсиями, арбовирусами и спирохетами наблюдается относительно безвредный комменсализм. В свою очередь еще неизвестные защитные реакции организма ведут к постеленному освобождению клеща от возбудителя или наблюдается пожизненное носительство последнего. Пребывание в организме на различных стадиях жизненного цикла может влиять не только на сроки переживания и интенсивность размножения возбудителей, но и вызывать изменения биологических свойств. Так, для воз-fiymrrei™ пятнистой лихорадки Скалистых гор описаны как обратимые изменения вирулентности под влиянием зимовки в клещах или их линьки, так и более сложные явления интерференции между штаммами возбудителя. Наблюдалась интерференция в клещах между вирусом колорадской клещевой лихорадки и риккетсиями-возбудителями пятнистой лихорадки Скалистых гор. При лабораторном заражении клещей Ornuhodoros papiUipes возбудителем клещевого спирохетоза н риккетсиями Бернета в их организме часто наблюдалось полное взаимоподавление обоих микроорганизмов. Напротив, в Ixodes persukatus
263
наблюдалось длительное и устойчивое существование риккетснй Бер-нета и вируса клещевого энцефалита.
Низкие температуры не приводят к необратимой инактивации или полной гибели возбудителей в клещах, что обеспечивает возможность их переэимования в организме переносчика в умеренных широтах. Питание на имунных хозяевах также не приводит к освобождению клещей от микроорганизмов, как это установлено в опытах с Coxiella burneti и Rickettsia sibiricus. В то же время в опытах с вирусом клещевого энцефалита питание иммунной кровью не влияло на его судьбу в организме нимф и имаго, вызывало нейтрализацию в личинках.
Многие экологические, морфологические и физиологические особенности иксодовых и аргасовых клещей исключительно благоприятны для биологической передачи возбудителей болезней. В то же время значительные промежутки времени между последовательными приемами пищи затрудняют механическую передачу возбудителей болезней. В большинстве случаев возбудители выделяются вместе со слюной, но для риккетснй и, возможно, бактерий важную роль приобретает выведение их из организма клеща вместе с фекалиями. Для риккетснй Берне! а контаминативный путь передачи от клещей теплокровным является, по-видимому, основным ввиду высокой инфекциознос-ти фекалий и многомесячного переживания в них возбудителя. Иноку-лятивной передаче возбудителей благоприятствует многодневное питание иксодовых клещей, сопровождающееся выделением в ранку большого количества слюны. В то же время в период питания выделяется большая масса фекалий и полупереваренной крови, что создает высокую концентрацию возбудителя не только на теле хозяина, но и на окружающих предметах.
Строение ротового аппарата у большинства видов клещей вызывает образование очага воспаления, заполненного кровью, обрывками тканей, тканевой жидкостью. Таким образом, клещи поглощают возбудителей не только с кровью, но и с лимфатической жидкостью и продуктами воспаления. Вместе с тем в рану выделяется слюна клеща: акты кровососания и слюноотделения чередуются, при этом выводятся избыточная вода и соли.
Клещи благодаря характеру внутриклеточного пищеварения
ПереПССлТ ЛЮиЫС иаКГСрпоЛЪНЫС ИНц^КЦйН, ПриТИВ ВОЗиуДНТёЛСи Ки-
торых в их организме нет или слабы специфические защитные факторы. Запрещенным фактором являются различные лизоцимы. Заражение Ixodes persulcatus вирусом клещевого энцефалита происходит чаще всего в первый момент поступления возбудителя в организм переносчика. Это связано, по-видимому, с тем, что секреторные альвеолы слюнных желез голодных особей недоразвиты и созревание их происходит уже на стадии питания. Возможно также, что на этой стадии доразвиваются и другие ткани, прежде всего пищеварительные.
264
Известно, что на первых этапах существования возбудителей в организме беспозвоночного важными компонентами внутрикишеч-ной среды являются ткани позвоночного, к которым присоединяется секрет слюнных желез, а в дальнейшем и пищеварительные соки, экскреты клеток кишечника. Таким образом, переход возбудителя из позвоночного в беспозвоночное происходит как бы смягченно благодаря довольно значительным остаткам тканей позвоночного хозяина. При обратном процессе (введение возбудителя в организм позвоночного) его сопровождает «остаток» среды беспозвоночного, иногда довольно значительный по объему, в виде слюны переносчика. Выделение слюны, содержащей возбудителей вирусной или риккетсиозной природы, чрезвычайно важно на последнем, завершающем этапе существования возбудителя в организме иксодового клеща. Повышение вероятности заражения позвоночного связано с простым количественным увеличением поступления инфекта в рану. Однако нельзя исключать и роль слюны переносчика, особенно учитывая что на более поздних этапах кровососания, когда передача наиболее вероятна, количество ее, выделяемое в ранку, растет.
Какой компонент играет в составе слюны роль адью ванта, скатать трудно, так как известно, что глико- и мукопротеиды слюны клещей или другие комплексы углеводов с белками являются антикоагулянтами — ингибиторами тромбокиназы и блокируют превращение протромбина в тромбин. Слюноотделение у аргасовых клещей не играет существенной роли при создании адьюваита для возбудителей. Эту функцию выполняет коксальная жидкость, являющаяся в большей или меньшей мере дериватом гемолимфы.
Клещи-краснотелки сосут не кровь, а тканевую жидкость с разрушенными под влиянием слюны мальпигиевьши клетками кожи теплокровного хозяина. Эта тканевая жидкость с жидкой фазой экссудата очага воспаления фагоцитируется пищеварительными клетками средней кишки тромбикулид. По-видимому, именно с этими особенностями питания связана высокая степень адаптации к организму краснотелок у возбудителей лихорадки пуцугамуши Rickettsia tsutsug-amushi, которые не размножаются в других клещах и не передаются ими трасфазово.
У тромбикулид фаза личинки является первой и единственной, на которой риккетсии могут быть восприняты и в ниыкуушщсм, дочернем, поколении переданы через укус. Белок, получаемый хищными нимфами и имаго тромбикулид за счет поглощенной гемолимфы насекомых, вернее, содержимого их яиц, способствует переживанию риккетсии в период трансфазовой и трансовариальной передачи.
Все это говорит о том, что кровососущие клещи являются биологическими переносчиками многих вирусных, риккетсиозных и бактериальных болезней человека и основными компонентами, биологическими факторами, природных очагов этих болезней.
265