

Билет 16.
Основные функции хлорофилла — поглощение света, превращение световой энергии в электронную и передача ее посредствомван-дер-ваальсового (диполь-дипольного) взаимодействия соседним молекулам. По цепи хлорофиллов электронная энергия передается к реакционному центру фотосинтеза, где используется для пространственного разделения заряда и последующих окислительно-восстановительных реакций. Хлорофиллы также входят в состав реакционных центров зеленых растений, где играют роль первичных доноров электронов.
В растениях хлорофиллы в составе антенн и реакционных центров расположены в фотосинтетических мембранах, где пространственно закрепляются в определенных местах с помощью фитильных боковых цепей и дополнительного комплексообразования между ионом Mg2+ и полипептидными цепями белков.
Билет 17
Фикобилины — красные и синие пигменты, содержащиеся у цианобактерий и некоторых водорослей. Исследования показали, что красные водоросли и цианобактерий наряду с хлорофиллом а содержат фикобилины. В основе химического строения фикобилинов лежат четыре пиррольные группировки. В отличие от хлорофилла у фикобилинов пиррольные группы расположены в виде открытой цепочки. Фикобилины представлены пигментами: фикоцианином, фикоэритрином и аллофикоцианином. Фикоэритрин — это окисленный фикоцианин. Красные водоросли в основном содержат фикоэритрин, а цианобактерий — фикоцианин. Фикобилины образуют прочные соединения с белками (фикобилинпротеиды). Связь между фикобилинами и белками разрушается только кислотой. Предполагается, что карбоксильные группы пигмента связываются с аминогруппами белка. Необходимо отметить, что в отличие от хлорофиллов и каротиноидов, расположенных в мембранах, фикобилины концентрируются в особых гранулах (фикобилисомах), тесно связанных с мембранами тилакоидов.

Фикобилины поглощают лучи в зеленой и желтой частях солнечного спектра. Это та часть спектра, которая находится между двумя основными линиями поглощения хлорофилла. Фикоэритрин поглощает лучи с длиной волны 495— 565 нм, а фикоцианин — 550— 615 нм. Сравнение спектров поглощения фикобилинов со спектральным составом света, в котором проходит фотосинтез у цианобактерий и красных водорослей, показывает, что они очень близки. Это позволяет считать, что фикобилины поглощают энергию света и, подобно каротиноидам, передают ее на молекулу хлорофилла, после чего она используется в процессе фотосинтеза. Наличие фикобилинов у водорослей является примером приспособления организмов в процессе эволюции к использованию участков солнечного спектра, которые проникают сквозь толщу морской воды (хроматическая адаптация). Как известно, красные лучи, соответствующие основной линии поглощения хлорофилла, поглощаются, проходя через толщу воды. Наиболее глубоко проникают зеленые лучи, которые поглощаются не хлорофиллом, а фикобилинами.
Билет 19.
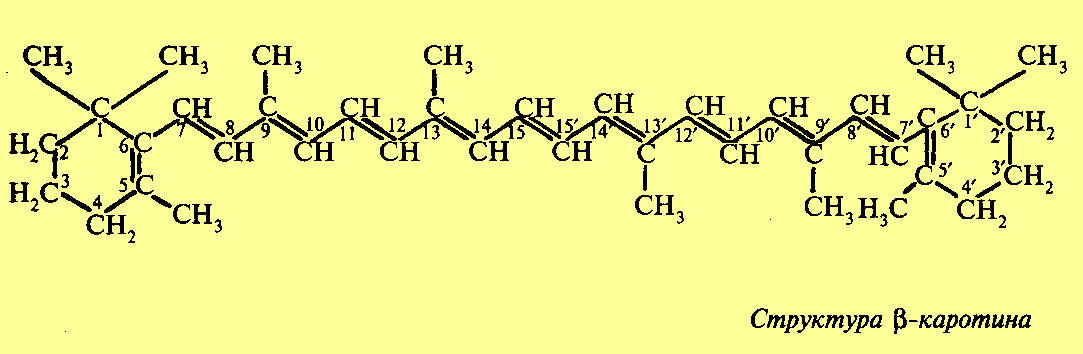
Наряду с зелеными пигментами в хлоропластах и хроматофорах содержатся пигменты, относящиеся к группе каротиноидов. Каротйнойды — это желтые и оранжевые пигменты алифатического строения, производные изопрена. Каротйнойды содержатся во всех высших растениях и у многих микроорганизмов. Это самые распространенные пигменты с разнообразными функциями. Каротинойды, содержащие кислород, получили название ксантофиллы. Основными представителями каротиноидов у высших растений являются два пигмента — каротин (оранжевый) и ксантофилл (желтый). Каротин состоит из 8 изопреновых остатков. При разрыве углеродной цепочки пополам и образовании на конце спиртовой группы каротин превращается в 2 молекулы витамина А. Обращает на себя внимание сходство в структуре фитоласпирта, входящего в состав хлорофилла, и углеродной цепочки, соединяющей пионовые кольца каротина. Предполагается, что фитол возникает как продукт гидрирования этой части молекулы каротиноидов. Поглощение света каротиноидами, их окраска, а также способность к окислительно-восстановительным реакциям обусловлены наличием конъюгированных двойных связей, b каротин имеет два максимума поглощения, соответствующие длинам волн 482 и 452 нм. В отличие от хлорофиллов каротиноиды не поглощают красные лучи, а также не обладают способностью к флуоресценции. Подобно хлорофиллу каротиноиды в хлоропластах и хроматофорах находятся в виде нерастворимых в воде комплексов с белками.
Физиологическая роль каротиноидов.
Уже тот факт, что каротиноиды всегда присутствуют в хлоропластах, позволяет считать, что они принимают участие в процессе фотосинтеза. Однако не отмечено ни одного случая, когда в отсутствие хлорофилла этот процесс осуществляется. В настоящее время установлено, что каротиноиды, поглощая определенные участки солнечного спектра, передают энергию этих лучей на молекулы хлорофилла. Тем самым они способствуют использованию лучей, которые хлорофиллом не поглощаются. Физиологическая роль каротиноидов не ограничивается их участием в передаче энергии на молекулы хлорофилла. По данным русского исследователя Д.И. Сапожникова, на свету происходит взаимопревращение ксантофиллов (виолаксантин превращается в зеаксантин), что сопровождается выделением кислорода. Спектр действия этой реакции совпадает со спектром поглощения хлорофилла, что позволило высказать предположение об ее участии в процессе разложения воды и выделения кислорода при фотосинтезе.
Имеются данные, что каротиноиды выполняют защитную функцию, предохраняя различные органические вещества, в первую очередь молекулы хлорофилла, от разрушения на свету в процессе фотоокисления. Опыты, проведенные на мутантах кукурузы и подсолнечника, показали, что они содержат протохлорофиллид (темновой предшественник хлорофилла), который на свету переходит в хлорофилл а, но разрушается. Последнее связано с отсутствием способности исследованных мутантов к образованию каротиноидов. Ряд исследователей указывают, что каротиноиды играют определенную роль в половом процессе у растений. Известно, что в период цветения высших растений содержание каротиноидов в листьях уменьшается. Одновременно оно заметно растет в пыльниках, а также в лепестках цветков. По мнению П. М. Жуковского, микроспорогенез тесно связан с метаболизмом каротиноидов. Незрелые пыльцевые зерна имеют белую окраску, а созревшая пыльца — желто-оранжевую. В половых клетках водорослей наблюдается дифференцированное распределение пигментов. Мужские гаметы имеют желтую окраску и содержат каротиноиды. Женские гаметы содержат хлорофилл. Высказывается мнение, что именно каротин обусловливает подвижность сперматозоидов. По данным В. Мевиуса, материнские клетки водоросли хламидомонады образуют половые клетки (гаметы) первоначально без жгутиков, в этот период они еще не могут передвигаться в воде. Жгутики образуются только после освещения гамет длинноволновыми лучами, которые улавливаются особым каротиноидом — кроцетином.
Образование каротиноидов.
Синтез каротиноидов не требует света. При формировании листьев каротиноиды образуются и накапливаются в пластидах еще в тот период, когда зачаток листа защищен в почке от действия света. В начале освещения образование хлорофилла в этиолированных проростках сопровождается временным падением содержания каротиноидов. Однако затем содержание каротиноидов восстанавливается и даже повышается с увеличением интенсивности освещения. Установлено, что между содержанием белка и каротиноидов имеется прямая коррелятивная связь. Потеря белка и каротиноидов в срезанных листьях идет параллельно. Образование каротиноидов зависит от источника азотного питания. Более благоприятные результаты по накоплению каротиноидов получены при выращивании растений на нитратном фоне по сравнению с аммиачным. Недостаток серы резко уменьшает содержание каротиноидов. Большое значение имеет соотношение — Ca/Mg в питательной среде. Относительное увеличение содержания кальция приводит к усиленному накоплению каротиноидов по сравнению с хлорофиллом. Противоположное влияние оказывает увеличение содержания магния.