

Билет 13.
Фотосинтез, образование зелеными растениями и некоторыми бактериями органических веществ с использованием энергии солнечного света. Происходит при участии пигментов (у растений хлорофиллов). В основе фотосинтеза лежатокислительно-восстановительные реакции, в которых электроны переносятся от донора (например, H2O, H2S) к акцептору (CO2) с образованием восстановленных соединений (углеводов) и выделением O2 (если донор электронов H2O), S (если донор электронов, например, H2S) и др.
В связи с тем, что углеводы составляют основную массу продуктов биосинтетической деятельности растений, химическое уравнение фотосинтеза обычно записывают в виде:
![]()
Для
этой реакции ![]() 469,3
кДж/моль, понижение энтропии 30,3
Дж/(К·моль),
469,3
кДж/моль, понижение энтропии 30,3
Дж/(К·моль), ![]() -479
кДж/моль. Квантовый расход фотосинтеза
для одноклеточных водорослей в
лабораторных условиях составляет 8-12
квантов на молекулуCO2.
Утилизация при фотосинтезе энергии
солнечного излучения, достигающего
земной поверхности, составляет не более
0,1% всей ФАР. Наиболее продуктивные
растения (например, сахарный тростник)
в среднем за год усваивают около 2%
энергии падающего излучения, а зерновые
культуры - до 1%. Обычно суммарная
продуктивность фотосинтеза ограничена
содержанием CO2 в
атмосфере (0,03-0,04% по объему), интенсивностью
света и температурой. Зрелые листья
шпината в атмосфере нормального состава
при 25 0C
на свету насыщающей интенсивности (при
солнечном освещении) дают несколько
литров O2 в
час на грамм хлорофилла или на
килограмм сухого веса. Для
водорослей Chlorella
pyrenoidosa при 35°C
повышение концентрации CO2 от
0,03 до 3% позволяет повысить выход O2 в
5 раз, такая активация является предельной.
-479
кДж/моль. Квантовый расход фотосинтеза
для одноклеточных водорослей в
лабораторных условиях составляет 8-12
квантов на молекулуCO2.
Утилизация при фотосинтезе энергии
солнечного излучения, достигающего
земной поверхности, составляет не более
0,1% всей ФАР. Наиболее продуктивные
растения (например, сахарный тростник)
в среднем за год усваивают около 2%
энергии падающего излучения, а зерновые
культуры - до 1%. Обычно суммарная
продуктивность фотосинтеза ограничена
содержанием CO2 в
атмосфере (0,03-0,04% по объему), интенсивностью
света и температурой. Зрелые листья
шпината в атмосфере нормального состава
при 25 0C
на свету насыщающей интенсивности (при
солнечном освещении) дают несколько
литров O2 в
час на грамм хлорофилла или на
килограмм сухого веса. Для
водорослей Chlorella
pyrenoidosa при 35°C
повышение концентрации CO2 от
0,03 до 3% позволяет повысить выход O2 в
5 раз, такая активация является предельной.
Бактериальный фотосинтез и общее ур-ние фотосинтеза.
Наряду с фотосинтезом высших растений и водорослей, сопровождаемым выделением O2, в природе осуществляется бактериальный фотосинтез, в к-ром окисляемым субстратом является не вода, а др. соединения, обладающие более выраженными восстановит. св-ми, напр. H2S, SO2. Кислород при бактериальном фотосинтезе не выделяется. Фотосинтезирующие бактерии способны использовать не только видимое, но и ближнее ИК излучение (до 1000 нм) в соответствии со спектрами поглощения преобладающих в них пигментов - бактериохлорофиллов. Бактериальный фотосинтез не имеет существенного значения в глобальном запасании солнечной энергии, но важен для понимания общих механизмов фотосинтеза. Обобщенное ур-ние фотосинтеза можно записать в виде:
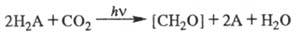
Билет 15.
Пигментные сист-мы.
Для того чтобы свет мог оказывать влияние на растительный организм и, в частности, быть использованным в процессе фотосинтеза, необходимо его поглощение фоторецепторами-пигментами. Пигменты — это окрашенные вещества. Пигменты поглощают свет определенной длины волны. Непоглощенные участки солнечного спектра отражаются, что и обусловливает окраску пигментов. Так, зеленый пигмент ХЛ поглощает красные и синие лучи, тогда как зеленые лучи в основном отражаются. Видимая часть солнечного спектра включает длины волн от 400 до 700 нм. Вещества, поглощающие весь видимый участок спектра, кажутся черными. Пигменты, сконцентрированные в пластидах, можно разделить на три группы: ХЛы, каротиноиды, фикобилины.
Источник энергии в фотосинтезе. Роль световых реакций фотосинтеза заключается в том, что в световую фазу синтезируются молекула АТФ и молекулы-переносчики протонов, то есть НАДФ Н2.
Окисл.-восстановительные реакции с участием ХЛа. В процессе фотосинтеза модификация ХЛа не происходит, он всего лишь помогает окислению донора и восстановлению акцептора, при участии кванта света. Когда hv поподает на молекулу ХЛа с него выбивается е` и восстанавливает акцептор, но за очень короткое время ХЛ оттягивает на себя е` с донора. Система сопряженных = связей в молекуле ХЛа обеспечивает потерю е`а при действии hv.