

23.Основные углеводы животных, их содержание в тканях, биологическая роль. Основные углеводы пищи. Переваривание углеводов
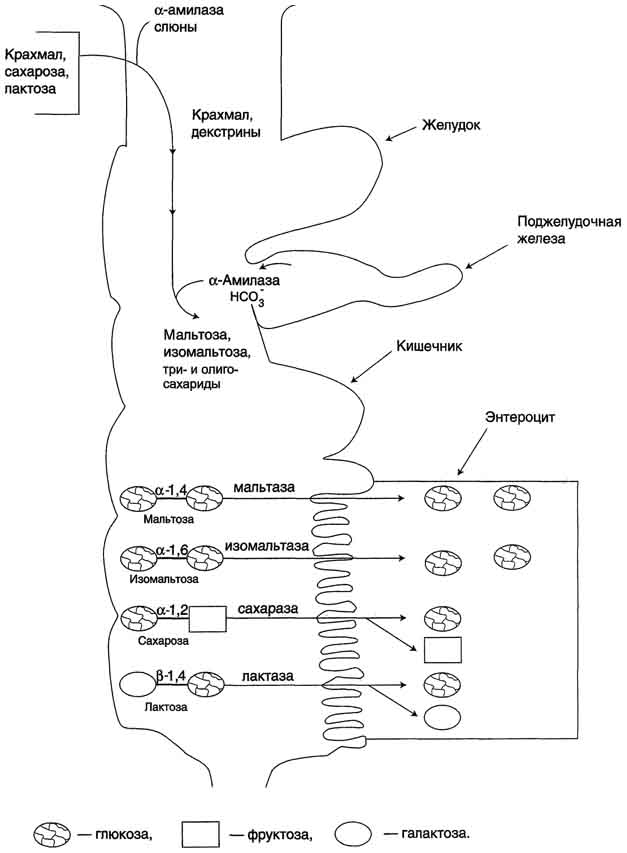
Гликоген – главный резервный полисахарид высших животных и человека, построенный из остатков D-глюкозы. Эмпирическая формулагликогена, как и крахмала, (С6Н10О5)n. Гликоген содержится практически во всех органах и тканях животных и человека; наибольшее количество обнаружено в печени и мышцах. Молекулярная масса гликогена 105–108 Да и более. Его молекула построена из ветвящихся полиглюкозидных цепей, в которых остатки глюкозы соединены α-1–>4-гликозидными связями. В точках ветвления имеются α-1–>6-гликозидные связи. По строению гликоген близок к амилопектину. В молекуле гликогена различают внутренние ветви – участки от периферической точки ветвления до нередуцирующего конца цепи. Гликоген характеризуется более разветвленной структурой, чем амилопектин; линейные отрезки в молекуле гликогена включают 11–18 остатков α-D-глюкопиранозы.
При гидролизе гликоген, подобно крахмалу, расщепляется с образованием сначала декстринов, затем мальтозы и, наконец, глюкозы.
Различают шесть основных классов гликозаминогликанов . Каждый из гликозаминогликанов содержит характерную для него повторяющуюся дисахаридную единицу; во всех случаях (кроме кератансульфатов) эта единица содержит либо глюкуроновую, либо идуроновую кислоту. Все гликозаминогликаны, за исключением гиалуроновой кислоты, содержат остатки моносахаридов с О- или N-сульфатной группой. Гликозаминогликаны значительно различаются по размерам, их молекулярные массы в пределах от 104 Да для гепарина до 107 Да для гиалуроновой кислоты.
Выделенные индивидуальные гликозаминогликаны могут содержать смесь цепей различной длины. Гликозаминогликаны как основное скрепляющее вещество связаны со структурными компонентами костей и соединительной ткани. Их функция состоит также в удержании большой массы воды и в заполнении межклеточного пространства. Иными словами, гликозаминогликаны – основной компонент внеклеточного вещества – жела-тинообразного вещества, заполняющего межклеточное пространство тканей. Они также содержатся в больших количествах в синовиальной жидкости – это вязкий материал, окружающий суставы, который служит смазкой и амортизатором. Поскольку водные растворы гликозаминогликанов гелеобразны, их называют мукополисахаридами.
Наконец, если цепи гликозаминогликана присоединены к белковой молекуле, соответствующее соединение называют протеогликаном.
Протеогликаны образуют основное вещество внеклеточного матрикса. В отличие от простых гликопротеинов, которые содержат только несколько процентов углеводов (по массе), протеогликаны могут содержать до 95% (и более) углеводов.
Крахмал - наиболее важный углеводный компонент пищевого рациона. Это резервный полисахарид растений, содержащийся в наибольшем количестве (до 45% от массы сухого вещества) в зёрнах злаков (пшеница, кукуруза, рис и др.), а также луковицах, стеблях и клубнях растений (в картофеле примерно 65%). Крахмал - разветвлённый полисахарид, состоящий из остатков глюкозы (гомогликан). Он находится в клетках растений в виде гранул, практически нерастворим в воде. Крахмал состоит из амилозы и амилопектина. Амилоза - неразветвлённый полисахарид, включающий 200-300 остатков глюкозы, связанных α-1,4-гликозидной связью. Благодаря α-конфигурации глюкозного остатка, полисахаридная цепь имеет конформацию спирали. Синяя окраска при добавлении йода к раствору крахмала обусловлена наличием такой спирали. Амилопектин имеет разветвлённую структуру. В местах ветвления остатки глюкозы соединены α-1,6-гликозидными связями. Линейные участки содержат примерно 20-25 остатков глюкозы. При этом формируется древовидная структура, в которой имеется лишь одна аномерная ОН-группа. Крахмал - высокомолекулярное соединение, включающее сотни тысяч остатков глюкозы. Его молекулярная масса составляет порядка 105-108 Д.
Биологическое значение углеводов:
Углеводы выполняют структурную функцию, то есть участвуют в построении различных клеточных структур (например, клеточных стенок растений).
Углеводы выполняют защитную роль у растений (клеточные стенки, состоящие из клеточных стенок мертвых клеток защитные образования — шипы, колючки и др.).
Углеводы выполняют пластическую функцию — хранятся в виде запаса питательных веществ, а также входят в состав сложных молекул (например, пентозы (рибоза идезоксирибоза) участвуют в построении АТФ, ДНК и РНК.
Углеводы являются основным энергетическим материалом. При окислении 1 грамма углеводов выделяются 4,1 ккал энергии и 0,4 г воды.
Углеводы участвуют в обеспечении осмотического давления и осморегуляции. Так, в крови содержится 100—110 мг/% глюкозы. От концентрации глюкозы зависитосмотическое давление крови.
Углеводы выполняют рецепторную функцию — многие олигосахариды входят в состав воспринимающей части клеточных рецепторов или молекул-лигандов.
24.См.
вопрос 22.
25Переваривание углеводов.
 26.
Глюкоза является
альдогексозой. Она может существовать
в линейной и циклической формах.
Циклическая форма глюкозы, предпочтительная
в термодинамическом отношении,
обусловливает химические свойства
глюкозы. Как и все гексозы, глюкоза имеет
4 асимметричных углеродных атома,
обусловливающих наличие сте-реоизомеров.
Возможно образование 16 стереоизомеров,
наиболее важные из которых D- и L-глюкоза.
Эти типы изомеров зеркально отображают
друг друга. Расположение Н- и ОН-групп
относительно пятого углеродного атома
определяет принадлежность глюкозы к
D- или L-ряду. В организме млекопитающих
моносахариды находятся в D-конфигурации,
так как к этой форме глюкозы специфичны
ферменты, катализирующие её превращения.
В растворе при образовании циклической
формы моносахарида образуются ещё 2
изомера (α- и β-изомеры), называемые
аномерами, обозначающие определённую
конформацию Н- и ОН-групп относительно
С, У α-D-глюкозы ОН-группа располагается
ниже плоскости кольца, а у β-D-глюкозы,
наоборот, над плоскостью кольца. Поскольку
в составе основных углеводов пищи
преобладает глюкоза, её можно считать
основным продуктом переваривания
углеводов. Другие моносахариды,
поступающие из кишечника в процессе
метаболизма, могут превращаться в
глюкозу или продукты её метаболизма.
Часть глюкозы в печени депонируется в
виде гликогена, а другая часть через
общий кровоток доставляется и используется
разными тканями и органами. При нормальном
рационе питания концентрация глюкозы
в крови поддерживается на уровне -3,3-5,5
ммоль/л (60-100 мг/дл). А в период пищеварения
её концентрация может повышаться
примерно до 150 мг/дл (8 ммоль/л).
26.
Глюкоза является
альдогексозой. Она может существовать
в линейной и циклической формах.
Циклическая форма глюкозы, предпочтительная
в термодинамическом отношении,
обусловливает химические свойства
глюкозы. Как и все гексозы, глюкоза имеет
4 асимметричных углеродных атома,
обусловливающих наличие сте-реоизомеров.
Возможно образование 16 стереоизомеров,
наиболее важные из которых D- и L-глюкоза.
Эти типы изомеров зеркально отображают
друг друга. Расположение Н- и ОН-групп
относительно пятого углеродного атома
определяет принадлежность глюкозы к
D- или L-ряду. В организме млекопитающих
моносахариды находятся в D-конфигурации,
так как к этой форме глюкозы специфичны
ферменты, катализирующие её превращения.
В растворе при образовании циклической
формы моносахарида образуются ещё 2
изомера (α- и β-изомеры), называемые
аномерами, обозначающие определённую
конформацию Н- и ОН-групп относительно
С, У α-D-глюкозы ОН-группа располагается
ниже плоскости кольца, а у β-D-глюкозы,
наоборот, над плоскостью кольца. Поскольку
в составе основных углеводов пищи
преобладает глюкоза, её можно считать
основным продуктом переваривания
углеводов. Другие моносахариды,
поступающие из кишечника в процессе
метаболизма, могут превращаться в
глюкозу или продукты её метаболизма.
Часть глюкозы в печени депонируется в
виде гликогена, а другая часть через
общий кровоток доставляется и используется
разными тканями и органами. При нормальном
рационе питания концентрация глюкозы
в крови поддерживается на уровне -3,3-5,5
ммоль/л (60-100 мг/дл). А в период пищеварения
её концентрация может повышаться
примерно до 150 мг/дл (8 ммоль/л).
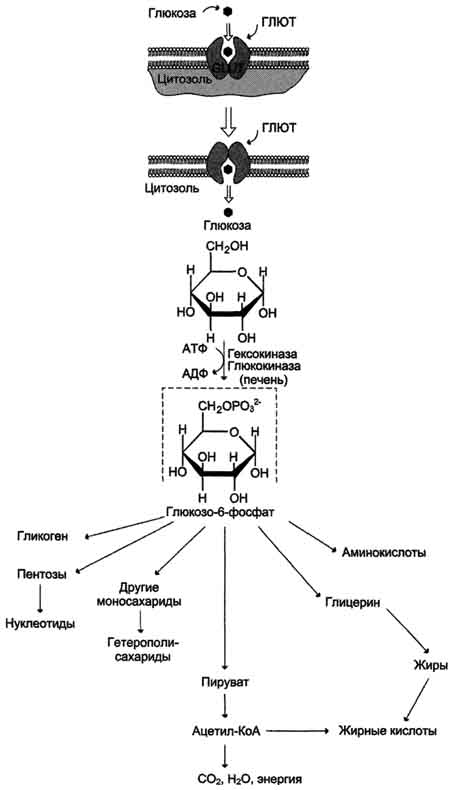
27. Источники и метаболизм глюкозы. Фософорилирование глюкозы В дальнейших превращениях в клетках глюкоза и другие моносахариды участвуют только в виде фосфорных эфиров. Фосфорилирование свободных моносахаридов - обязательная реакция на пути их использования, она приводит к образованию более реакционно-способных соединений и поэтому может рассматриваться как реакция активации. Глюкоза, поступающая в клетки органов и тканей, сразу же подвергается фосфорилированию с использованием АТФ. Эту реакцию во многих тканях катализирует фермент гексокиназа, а в печени и поджелудочной железе - фермент глюкокиназа. Фосфорилирование глюкозы - практически необратимая реакция, так как она протекает с использованием значительного количества энергии. Образование глюкозо-6-фосфата в клетке - своеобразная "ловушка" для глюкозы, так как мембрана клетки непроницаема для фосфорилированной глюкозы (нет соответствующих транспортных белков). Кроме того, Фосфорилирование уменьшает концентрацию свободной глюкозы в цитоплазме. В результате создаются благоприятные условия для облегчённой диффузии глюкозы в клетки из крови.
Глюкокиназа. Фосфорилирование глюкозы в гепатоцитах в период пищеварения обеспечивается свойствами глюкокиназы, которая имеет высокое значение Кm - 10 ммоль/л. В этот период концентрация глюкозы в воротной вене больше, чем в других отделах кровяного русла и может превышать 10 ммоль/л, а следовательно, активность глюкокиназы в гепатоцитах повышается. Следует отметить, что активность глюкокиназы, в отличие от гексокиназы, не ингибируется продуктом катализируемой реакции - глюкозо-6-фосфатом. Это обстоятельство обеспечивает повышение концентрации глюкозы в клетке в фосфорилированной форме, соответственно её уровню в крови. Как уже упоминалось, глюкоза проникает в гепатоциты путём облегчённой диффузии при участии транспортёра ГЛЮТ-2 (независимого от инсулина). ГЛЮТ-2, так же, как глюкокиназа, имеет высокую Кm, что способствует повышению скорости поступления глюкозы в гепатоциты в период пищеварения, следовательно, ускоряет её фосфорилирование и дальнейшее использование для депонирования. Хотя инсулин и не влияет на транспорт глюкозы, он усиливает приток глюкозы в гепатоциты в период пищеварения косвенным путём, индуцируя синтез глюкокиназы и ускоряя тем самым Фосфорилирование глюкозы. Преимущественное потребление глюкозы гепатоцитами, обусловленное свойствами глюкокиназы, предотвращает чрезмерное повышение её концентрации в крови в абсорбтивном периоде. Это, в свою очередь, снижает последствия протекания нежелательных реакций с участием глюкозы, например гликозилирования белков.
Гексокиназа отличается от глюкокиназы высоким сродством к глюкозе (Кm <0,1 ммоль/л). Следовательно, этот фермент, в отличие от глюкокиназы, активен при низкой концентрации глюкозы в крови, что характерно для постабсорбтивного состояния. Печень в этот период поглощает гораздо меньше глюкозы, так как скорость её внутриклеточного фосфорилирования глюкокиназой резко снижается. Тогда как потребление глюкозы мозгом, эритроцитами и другими тканями обеспечивается активной в этих условиях гексокиназой. Фермент гексокиназа может катализировать фосфорилирование не только D-глюкозы, но и других гексоз, хотя и с меньшей скоростью. Активность гексокиназы изменяется в зависимости от потребностей клетки в энергии. В качестве регуляторов выступают соотношение АТФ/АДФ и внутриклеточный уровень глюкозо-6-фосфата (продукта катализируемой реакции). При снижении расхода энергии в клетке повышается уровень АТФ (относительно АДФ) и глюкозо-6-фосфата. В этом случае активность гексокиназы снижается, и, следовательно, уменьшается скорость поступления глюкозы в клетку. Следует отметить, что в разных тканях гексокиназа присутствует в различных изоформах, отличающихся величиной Кm. Глюкокиназа печени (и почек) является изоформой IV (гексокиназа IV). В клетках мышц содержится гексокиназа II, а в клетках опухолевых тканей преобладает гексокиназа III, с более высоким, чем у гексокиназы II, сродством к глюкозе.
Дефосфорилирование глюкозо-6-фосфата Превращение глюкозо-6-фосфата в глюкозу возможно в печени, почках и клетках эпителия кишечника. В клетках этих органов имеется фермент глюкозо-6-фосфатаза, катализирующая отщепление фосфатной группы гидролитическим путём:
Глюкозо-6-фосфат +Н2О → Глюкоза + Н3РО4
Образовавшаяся свободная глюкоза способна диффундировать из этих органов в кровь. В других органах и тканях глюкозо-6-фосфатазы нет, и поэтому дефосфорилирование глюкозо-6-фосфата невозможно. Пример подобного необратимого проникновения глюкозы в клетку - мышцы, где глюкозо-6-фосфат может использоваться только в метаболизме этой клетки.
Метаболизм глюкозо-6-фосфата Глюкозо-6-фосфат может использоваться в клетке в различных превращениях, основными из которых являются: синтез гликогена, катаболизм с образованием СО2 и Н2О или лактата, синтез пентоз. Распад глюкозы до конечных продуктов служит источником энергии для организма. Вместе с тем в процессе метаболизма глюкозо-6-фосфата образуются промежуточные продукты, используемые в дальнейшем для синтеза аминокислот, нуклеотидов, глицерина и жирных кислот. Таким образом, глюкозо-6-фосфат - не только субстрат для окисления, но и строительный материал для синтеза новых соединений
 .
.
28. Катаболизм глюкозы - основной поставщик энергии для процессов жизнедеятельности организма. Окисление глюкозы до СО2 и Н2О (аэробный распад). Аэробный распад глюкозы можно выразить суммарным уравнением:
С6Н12О6 + 6 О2 → 6 СО2 + Н2О + 2820 кДж/моль.
Аэробный гликолиз
Аэробным гликолизом называют процесс окисления глюкозы до пировиноградной кислоты, протекающий в присутствии кислорода. Все ферменты, катализирующие реакции этого процесса, локализованы в цитозоле клетки.