

3 Экологическое значение углекислого газа воздуха, поступление со2 в растение и его регуляция
Экологическое значение углекислого газа воздуха:
– растения строят органические вещества своего тела за счет его ассимиляции при фотосинтезе. Для образования 1 г глюкозы необходимо поглотить 1,47 г (или 0,75 л) СО2 (учитывая его концентрацию, такое количество СО2 извлекается из 2500 л воздуха).
– Углекислый газ имеет большое значение и в химическом изменении минеральной части почв. Почвенный раствор, насыщенный углекислотой, растворяет многие соединения почвы, в частности кальцит (СаСО3), доломит (CaCО3 ∙ MgCО3), магнезит (MgCО3), сидерит (FeCО3).
Содержание углекислого газа в воздухе и его колебания
Относительное постоянство содержания углекислоты в атмосфере поддерживается благодаря равновесию всех составляющих биогеохимического круговорота углерода и буферной роли Мирового океана. Однако концентрация С02 в воздухе постоянно увеличивается, возможно, благодаря интенсивному поступлению в атмосферу углерода, выведенного из круговорота в прежние геологические эпохи, прежде всего, в результате интенсивного сжигания ископаемого топлива.
Намного больше углерода содержится в виде карбонатов в осадочных породах, живых и отмерших организмах, и даже в Мировом океане его в десятки раз больше, чем в атмосфере, что связано с высокой растворимостью СО2 в воде.
Расчеты показывают, что к 2035 г. содержание СО2 в атмосфере может удвоиться и составить около 0,06%. Следствием этого может быть глобальное потепление климата, обусловленное так называемым «тепличным эффектом». Его связывают с тем, что углекислый газ «прозрачен» для основной части солнечного света, но задерживает тепловое (инфракрасное) излучение от нагретой Солнцем поверхности Земли. По некоторым расчетам, увеличение концентрации СО2 в атмосфере в 2 раза может привести к повышению температуры поверхности Земли на 2-3°, причем оно будет минимальным в тропической зоне и максимальным (8-11°) в высоких широтах.
Сезонные колебания концентрации атмосферной углекислоты связаны с фотосинтезом и изменениями увлажнения, аэрации и температуры почвы, влияющими на почвенную микрофлору. Так, в высоких широтах после продолжительных дождей активность аэробных почвенных микроорганизмов падает, а содержание углекислоты в приземном слое воздуха снижается. В аридных областях угнетение почвенной микрофлоры и уменьшение содержания углекислоты в приземном слое воздуха вызывается засухой. Годичные колебания СО2 в воздухе особенно значительны в Северном полушарии, где преобладает суша. Весной в северных областях потребление углекислого газа развивающимися растениями значительно превышает его поступление из почвы.
Суточные колебания содержания углекислоты более выражены, чем сезонные. В фитоценозах обычно наблюдается связанное с фотосинтезом дневное уменьшение концентрации СО2. Например, в лесу начало фотосинтеза утром обычно приводит к резкому падению его содержания. К полудню температура повышается, дыхание усиливается и количество углекислоты в кронах отчасти выравнивается. После захода солнца фотосинтез прекращается, начинает преобладать дыхание и концентрация СО2 вблизи почвы повышается.
Изменение содержания углекислого газа происходит также при подъеме в горы. Например, на Памире на высоте 3800 м концентрация СО2 всего 0,012—0,020%.
Источники атмосферной углекислоты
Углекислота выделяется при извержениях вулканов (65 млрд т в год), сжигании горючих веществ, и некоторую прибавку дает разложение карбонатных горных пород. Однако главным источником углекислого газа в сообществах суши является дыхание почвы. Под ним понимают выделение СО2 эдафоном (совокупностью живых организмов почвы) в прилегающий к почве слой воздуха. В этом процессе на долю дыхания корней приходится до 10-30%, дыхание крупных животных очень мало, и основное количество СО2 образуется за счет микробиологической переработки остатков отмерших организмов (поэтому дыхание почвы является хорошим показателем интенсивности минерализации органического вещества почвы).
Поступление углекислоты в растение
Газообмен растения происходит путем диффузии, и на пути от атмосферного воздуха до хлоропластов двуокись углерода преодолевает ряд препятствий, определяющих в сумме диффузное сопротивление. В целом сопротивление транспорту углекислоты включает сопротивление пограничного слоя (воздушной пленки около листа), физиологически регулируемое устьичное сопротивление, диффузное сопротивление в системе межклетников, сопротивление при растворении и транспортировке СО2 в жидкой фазе клеточной стенки и в протоплазме, а также сопротивление в процессе карбоксилирования. Сопротивление пограничного слоя ниже, чем последующие устьичное и мезофильное сопротивления.
У наземных растений СО2 проникает в лист через устьица (прохождение через клетки эпидермиса возможно только при очень высоких его концентрациях, в естественных условиях не встречающихся). Устьичное сопротивление – главное препятствие для свободного поступления углекислоты в растение. Закрытые устьица практически полностью перекрывают поток СО2 в лист. При недостатке света, когда устьичные щели суживаются, устьичное сопротивление становится фактором, ограничивающим диффузию СО2. При широко раскрытых устьицах это сопротивление ниже сопротивления мезофилла.
Прошедшая через устьичную щель углекислота попадает в межклетники, где пополняется СО2, выделившимся при дыхании клеток. Сопротивление диффузии в системе межклетников зависит от строения листа. Обычно газопроводность теневых листьев выше, чем световых, а в мягких листьях она лучше, чем в жестких фотосинтезирующих органах.
В клеточных стенках углекислый газ растворяется, и этот процесс тормозит диффузию. Внутри клетки растворенная углекислота очень медленно (по сравнению с газообразной фазой) движется к хлоропластам, где присоединяется к акцептору. Карбоксилирование может отставать от световых реакций фотосинтеза и лимитировать скорость этого процесса в целом. Сопротивление карбоксилирования зависит от эффективности работы ферментов – карбоксилаз, отличающихся в разных группах растений. Так как ФЕП-карбоксилаза катализирует связывание углекислоты намного эффективнее, чем РДФ-карбоксилаза, сопротивление карбоксилирования у С4-видов значительно меньше, чем у С3-растений.
Устьичная регуляция поступления СО2
Изменяя ширину устьичных щелей, растение контролирует поступление углекислоты и одновременно идущую потерю воды. Количество, распределение, размеры, форма, подвижность устьиц видоспецифичны, но в некоторой степени изменчивы в зависимости от условий местообитаний и индивидуальных особенностей растений. Для поступления углекислого газа в растение решающее значение имеет площадь отверстий устьиц. С ее уменьшением устьичное сопротивление диффузии возрастает по экспоненте. У большинства растений от верстия устьиц занимают 0,5-1,5% поверхности листа.
Максимальная диффузия газов в лист достигается при максимальном открытии устьиц, которое зависит от формы замыкающих клеток и особенностей строения их оболочек. Степень открытости устьиц зависит от состояния растения и внешней среды. Из экологических факторов наиболее значимы свет, температура, влажность воздуха и водообеспеченность. Из внутренних факторов сильнее всего влияют на поступление углекислоты парциальное давление в межклетниках, гидратация растения, баланс ионов и фитогормонов. Из гормонов гибберелиновая кислота и цитокинины способствуют открыванию устьиц, а абсцизовая кислота – закрыванию. На состояние устьиц также влияют возраст листьев, стадия развития растения и эндогенные суточные ритмы.
При хорошем водоснабжении устьица открываются тем шире, чем интенсивнее освещение – фотоактивное открывание. Оно определяется не только прямым, но и косвенным действием света (через расход СО2 на свету при фотосинтезе). Хемоактивная регуляции устьичных движений связана с содержанием углекислоты в воздухе. Так, при снижении ее содержания ниже критического значения они открываются и в темноте. Реакцию устьиц индуцируют и другие вещества (SО2, озон, сульфат алюминия и др.). На состояние устьиц влияют также некоторые токсины паразитов (например, выделения фитопатогенного гриба Helminthosporium maydis), а также вещества, применяемые для борьбы с ними (например, фунгицид фенилмеркурацетат).
Температура влияет на скорость открывания устьиц через выработку энергии для их движения. При более высокой (но неэкстремальной) температуре энергии у клетки больше и устьица открываются быстрее. При сверхоптимальных температурах устьица также открываются не полностью, однако в жару (+40-45°) у некоторых видов ширина устьиц вновь достигает максимума.
Важный фактор регуляции состояния устьиц – водообеспеченность. Устьичная щель закрывается при потере воды и падении тургора замыкающих клеток. В сухом воздухе реакция закрывания устьиц наступит раньше, чем растение начнет испытывать серьезный недостаток воды. Ветер, сдувающий водяной пар с листа, усиливает этот эффект.
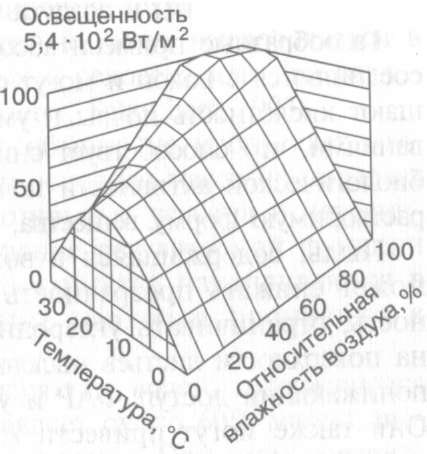 Совокупное
влияние внешних факторов обычно
обусловливает не максимальную,
а среднюю степень открытия устьиц.
Полное закрытие устьичных щелей вызывают
тоже только экстремально неблагоприятные
условия.
Совокупное
влияние внешних факторов обычно
обусловливает не максимальную,
а среднюю степень открытия устьиц.
Полное закрытие устьичных щелей вызывают
тоже только экстремально неблагоприятные
условия.
Физиологический механизм движений замыкающих клеток связан с изменением тургора. В расслабленном состоянии устьичные щели закрыты, и их открывание требует затрат энергии. Тургор может быть повышен активным транспортированием ионов (К+, РО43-), изменением проницаемости и ферментативными процессами. При этом существенны запас АТФ, процессы карбоксилирования, ферментативные превращения крахмала и сахаров.