

Униполярный и биполярный методы регистрации потенциала действия нервного ствола
При внеклеточном отведении от нервного ствола регистрируют не изменение потенциала на мембране, а падение напряжения на внеклеточном межэлектродном участке, вызванное протеканием локальных токов вдоль наружной поверхности аксонов от покоящихся участков к активному. С этим связан рост амплитуды регистрируемого внеклеточного потенциала действия (ПД) при увеличении числа возбужденных аксонов. Характер кривой ПД нерва во многом определяется условиями его отведения. Известны два основных способа отведения ПД нерва:
1) если оба электрода располагаются на неповрежденной поверхности нерва (биполярное отведение), то в этом случае ПД регистрируется в виде двухфазного колебания (рис. 89, 1);
2) если один из отводящих электродов располагают на неповрежденном участке нерва, а другой – на умерщвленном участке или на какой-либо неактивной ткани (униполярное отведение).
Регистрируемый при униполярном отведении ПД имеет вид однофазного колебания (рис. 89, 2).
Представленная на рис. 89 схема поясняет механизм возникновения двух- и однофазного ПД. Под влиянием раздражения (рис. 89, А), наносимого с помощью стимулирующих электродов (СЭ), в нерве возникает возбуждение (деполяризация). Перемещаясь вдоль нерва, оно достигает первого из отводящих электродов (ОЭ1). Так как возбужденный участок по отношению к невозбужденному электроотрицателен, а также вследствие того, что второй из отводящих электродов (ОЭ2) расположен на покоящемся участке, между электродами ОЭ1 и
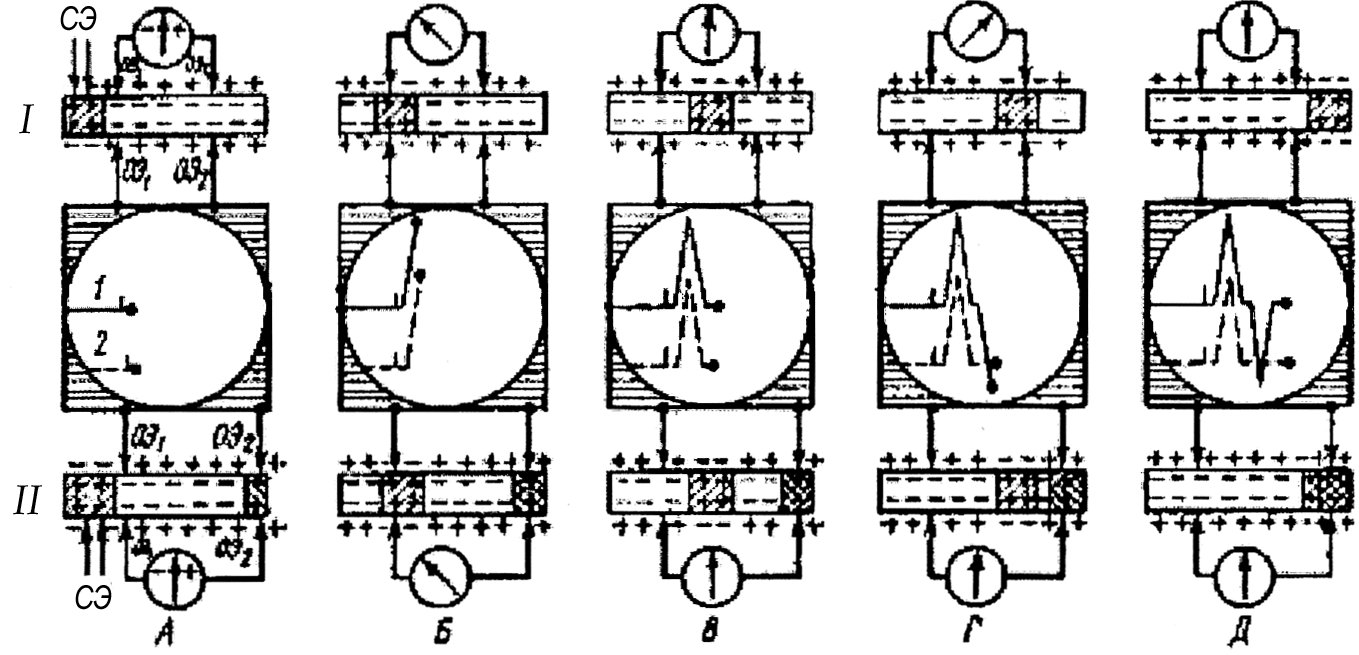
Рис. 89. Схема формирования кривой потенциала действия нерва
в непроводящей среде при биполярном (I) и униполярном (II) способах
отведения (пояснения в тексте)
и ОЭ2 возникает разность потенциалов. Под ее влиянием луч осциллографа будет отклоняться, регистрируя отрицательное колебание потенциала (рис. 89, Б). В зависимости от того, как подключены отводящие электроды к входу усилителя осциллографа, это колебание может быть направлено вверх или вниз от изолинии.
Принято так подключать отводящие электроды, чтобы отрицательное колебание отклоняло луч вверх. Если к электродам подключен гальванометр, то в нем произойдет отклонение стрелки влево. При дальнейшем движении ПД возбужденный участок будет находиться между электродами, т. е. разности потенциалов между ОЭ1 и ОЭ2 не будет, в результате этого луч осциллографа вернется к изолинии (рис. 89, В). Когда возбужденный участок достигнет ОЭ2, этот электрод становится отрицательным к ОЭ1, что вызовет отклонение луча осциллографа вниз (рис. 89, Г). Когда возбужденный участок минует электрод ОЭ2, разности потенциалов между ОЭ2 и ОЭ1 не будет, и луч осциллографа вернется к изолинии (рис. 89, Д). Так формируется двухфазный потенциал действия.
При униполярном отведении возбуждение под второй электрод не проводится (нерв под ним умерщвлен или проведение блокировано) – потенциал действия представлен однофазным отклонением луча осциллографа (см. рис. 89, 2).
Приведенные схемы отражают механизм формирования потенциалов, когда нерв помещен в непроводящую среду. В естественных условиях, когда нерв окружен тканями, и в искусственных – при помещении его в электролит – процессы формирования потенциалов протекают более сложно.
На рис. 90 приведены этапы формирования кривой ПД при униполярном способе отведения для тех случаев, когда нерв находится в проводящей среде, например во влажной камере. Индифферентный электрод располагают в отдаленной точке проводящей среды или помещают на неактивной ткани. При отведении в таких случаях приходится учитывать ряд факторов, не играющих роли при условии, что нерв окружен непроводящей средой:
1. Распространение потенциала по нерву связано не только с передвижением ионов через мембрану в отдельных волокнах, но и с перемещением их от неактивных участков к активным и обратно (анионы двигаются к покоящимся участкам, катионы – к возбужденным).
2. Распространение потенциала сопровождается возникновением местных токов между нервом и окружающими тканями. В фазу деполяризации мембран токи текут и по направлению к нерву, а в фазу реполяризации – в обратном направлении. Проходя через окружающие ткани, они вызывают появление вторичных полюсов, которые, в свою очередь, влияют на динамику местных потенциалов и на характер электрического поля, возникающего вокруг нерва.
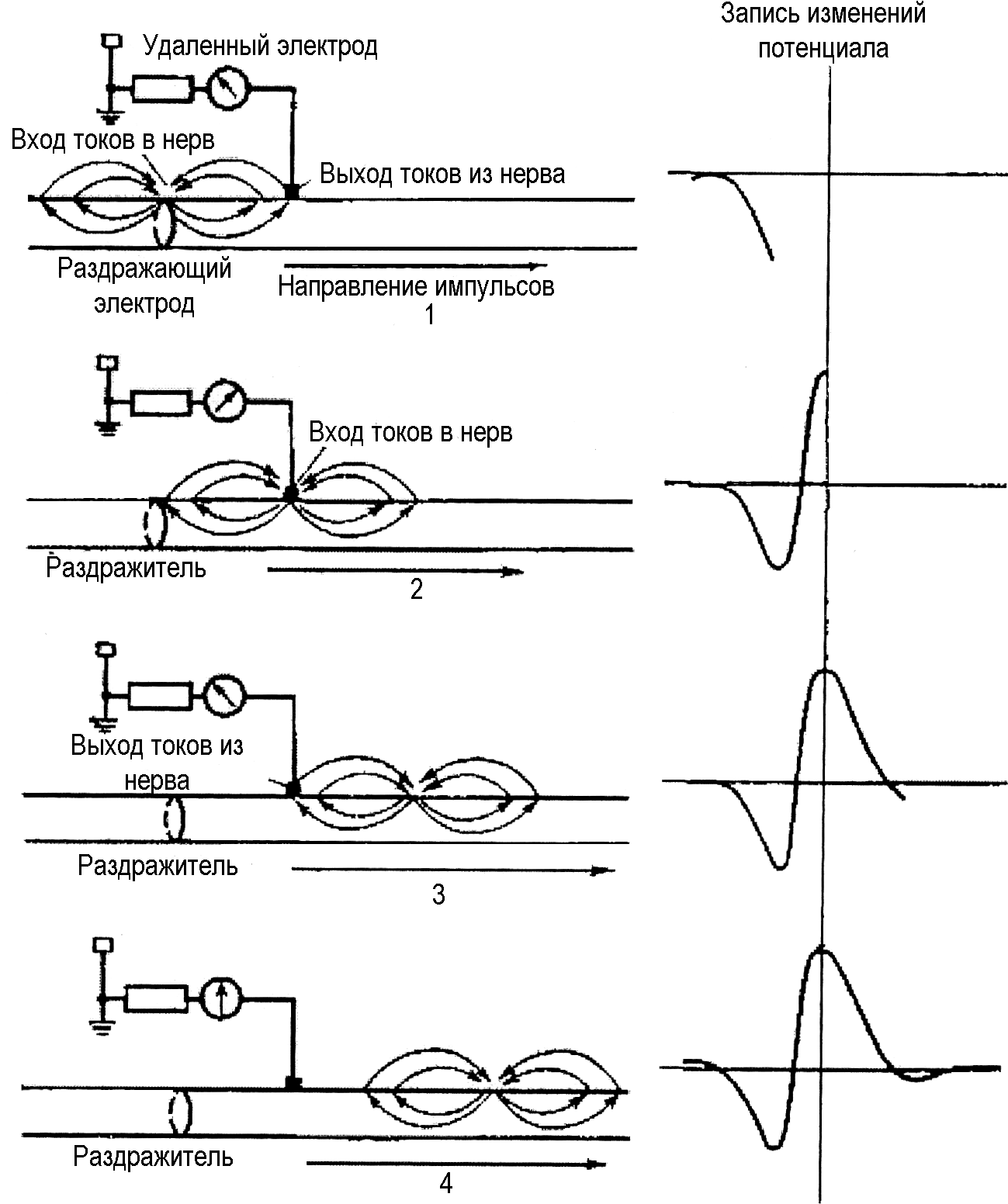
Рис. 90. Этапы формирования кривой потенциала действия нерва
(в проводящей среде или при отведении in situ)