НУКЛЕОТИДЫ, НУКЛЕОЗИДЫ, НУКЛЕИНОВЫЕ КИСЛОТЫ |
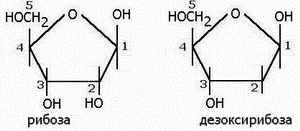
При полном гидролизе нуклеиновых кислот (НК) выделены соединения: а) пуриновые и пиримидиновые основания; б) моносахариды РИБОЗА или ДЕЗОКСИРИБОЗА (С5Н10О5); в) фосфорная кислота.
Строение моносахаридов:
Строение пуриновых оснований:
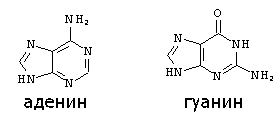
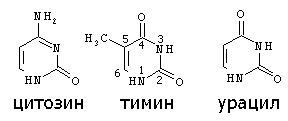
Строение пиримидиновых оснований:
Нуклеиновые кислоты имеют различающийся состав. В частности, дезоксирибонуклеиновые кислоты (ДНК) содержат дезоксирибозу, а рибонуклеиновые кислоты (РНК)- рибозу. Эти и другие отличия в составе нуклеиновых кислот приведены в таблице:
Одинаковые компоненты |
Отличающиеся компоненты |
|
ДНК |
РНК |
|
АДЕНИН ГУАНИН ЦИТОЗИН |
ДЕЗОКСИРИБОЗА ТИМИН |
РИБОЗА УРАЦИЛ |
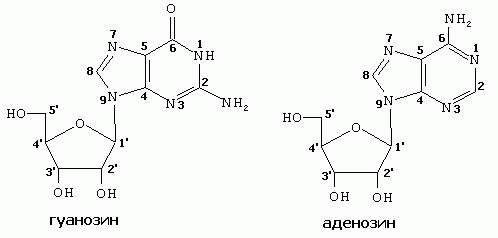
НУКЛЕОЗИДЫ- соединения азотистого основания и углеводов (рибозы и дезоксирибозы). Нуклеозиды образуются за счет N-гликозидной связи между ДЕВЯТЫМ атомом азота у ПУРИНОВЫХ (первым атомом азота- у ПИРИМИДИНОВЫХ) оснований и гидроксилом ПЕРВОГО атома углерода рибозы или дезоксирибозы. Во избежание путаницы, нумерация атомов азотистых оснований осуществляется арабскими цифрами, а у атомов углерода рибоз- арабскими цифрами со “штрихом”.
пуриновые |
|
пиримидиновые |
|
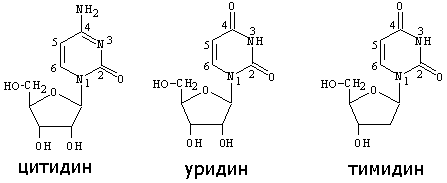
НУКЛЕОТИДЫ отличаются от нуклеозидов наличием остатков фосфорной кислоты (от одного до трех), связанных простой эфирной связью с гидроксилом 5’ атома углерода рибоз. Остатки фосфорных кислот между собой также связаны простой эфирной связью. В зависимости от числа остатков фосфорной кислоты в нуклеотидах различают МОНО-, ДИ- и ТРИФОСФОНУКЛЕОТИДЫ. Их номенклатура приведена в таблице:
азотистые основания |
нуклеозиды |
Нуклеотиды |
|
полное название |
Сокращенное |
||
аденин |
аденозин |
Аденозинмонофосфат |
АМФ |
гуанин |
гуанозин |
Гуанозинмонофосфат |
ГМФ |
цитозин |
цитидин |
Цитидинмонофосфат |
ЦМФ |
урацил |
уридин |
Уридинмонофосфат |
УМФ |
тимин |
тимидин |
тимидинмонофосфат |
ТМФ |
|
Собственно НУКЛЕИНОВЫЕ кислоты представляют собой полинуклеотидмонофосфаты. Полимерная цепь образуется за счет фосфодиэфирной связи между 3’- гидроксилом одного нуклеотида и 5’- гидроксилом другого. Таким образом, первичная структура нуклеиновых кислот представляет собой ПОРЯДОК чередования нуклеотидов в полинуклеотидной цепи. Один из концов этой цепи (изображаемый слева) имеет свободный гидроксил при 5’- атоме С, а другой (изображаемый справа)- свободный гидроксил при 3’- атоме углерода рибоз. Поскольку основой нуклеиновых кислот является САХАРОФОСФАТНЫЙ ОСТОВ, в сокращенных написаниях участков цепи используют лишь ОДНОБУКВЕННЫЕ СИМВОЛЫ соответствующего азотистого основания. Полное и схематичное обозначение участка полинуклеотидной цепи приведены ниже: 5’-НО-G-A-A-T-C-T-A-C-A-…3' Вследствие наличия сильнодиссоциирующих фосфатных групп, нуклеиновые кислоты (НК) легко образуют связи с оснОвными белками (высокое значение pI) с образованием НУКЛЕОПРОТЕИНОВ. Протеины отделяются от НК детергентами или после расщепления белков протеиназами НК осаждаются спиртом. |
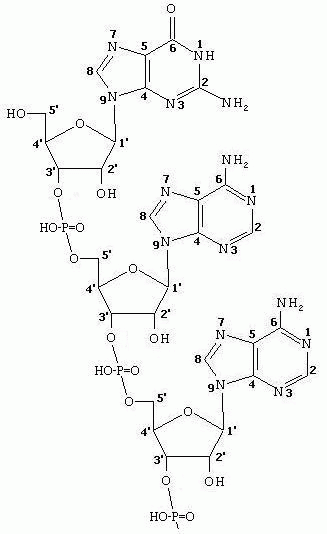
Подобно белкам, ДНК имеют первичную, вторичную и третичную структуру.
ПЕРВИЧНАЯ СТРУКТУРА есть последовательность чередования нуклеотидов в цепи ДНК и РНК. Сложность расшифровки структуры связана с наличием всего 4-х видов нуклеотидов при их огромном числе в молекуле.
ПРАВИЛА ЧАРГАФФА
1) сумма ПУРИНОВЫХ оснований в молекуле ДНК всегда равна сумме ПИРИМИДИНОВЫХ оснований.
2) А=Т, А/Т= 1; 3) Г=Ц, Г/Ц=1; 4) А+Т=Г+Ц; 5) если А+Т>Г+Ц, то АТ-тип.
Вторичная структура нуклеиновых кислот
Расшифрована впервые в 1953 г. Уотсоном и Крик (Watson, Crick).
Вторичная структура ДНК представляет собой свернутые в спираль ДВЕ комплементарно взаимодействующие и антипараллельные полинуклеотидные цепи. Образование вторичной структуры НК возможно вследствие проявления эффектов КОМПЛЕМЕНТАРНОСТИ и СТЭКИНГ-ВЗАИМОДЕЙСТВИЙ. Очень часто наблюдаются двунитевые спирализованные молекулы ДНК, замкнутые в кольцо с ковалентно связанными концами. Они не имеют разрывов у каждой в отдельности полинуклеотидной цепи. Подобные кольцевые ДНК, как правило, СУПЕРСПИРАЛИЗОВАНЫ, то есть кольцо дополнительно закручено в спираль. Суперспирализация- ПРАВИЛО, а не ИСКЛЮЧЕНИЕ, при условии отсутствия разрывов в фосфодиэфирных связях полинуклеотидной цепи.
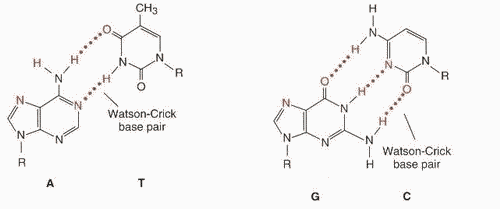
КОМПЛЕМЕНТАРНОСТЬ
- последовательность нуклеотидов в
одной цепи автоматически определяет
строго соответствующую ей последовательность
нуклеотидов в КОМПЛЕМЕНТАРНОЙ ей цепи.
Так, азотистое основание Аденин (А)
всегда взаимодействует только с
комплементарным ему азотистым основанием
Тимин (Т) в молекулах ДНК.
Одновременно
азотистые основания Гуанин (Г) одной
цепи взаимодействует только с
комплементарними им азотистыми
основаниями Цитозин (Ц) другой цепи (как
в ДНК, так и в РНК). Комплементарность
оснований обеспечивается системой
водородных связей. В молекулах РНК,
имеющих, в основном, однонитевую
структуру, на отдельных участках,
азотистые основания А взаимодействуют
с комплементарными им азотистыми
основаниями Урацил (У).
 Аналогично
происходит взаимодействие в процессах
транскрипции, когда на матрице ДНК
синтезируется молекула РНК (матричная,
транспортная и т.д.), и наоборот, когда
при участии РЕВЕРС-ТРАНСКРИПТАЗЫ
происходит синтез кДНК на матрице РНК.
Аналогично
происходит взаимодействие в процессах
транскрипции, когда на матрице ДНК
синтезируется молекула РНК (матричная,
транспортная и т.д.), и наоборот, когда
при участии РЕВЕРС-ТРАНСКРИПТАЗЫ
происходит синтез кДНК на матрице РНК.
СТЭКИНГ-ВЗАИМОДЕЙСТВИЯ- особого рода (Ван-дер-Ваальсовы) взаимодействия между выложенными в стопку (как монеты) друг над другом азотистых оснований.
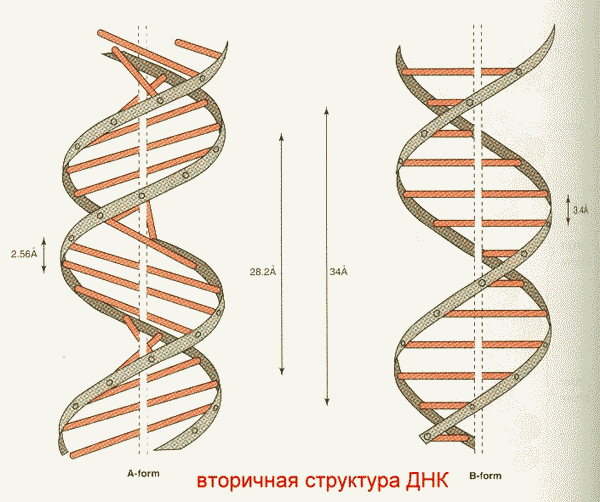
Имеются А, В, С и Z-формы двунитевых участков ДНК, отличающиеся наклонами плоскостей азотистых оснований друг относительно друга ( у А- 20о, В- 0о, С- 5о, Z- особая ломанная форма).
SBS (side-by-side)- форма (бок о бок). Две цепи расположены прямо и незакручены.
Третичная структура днк
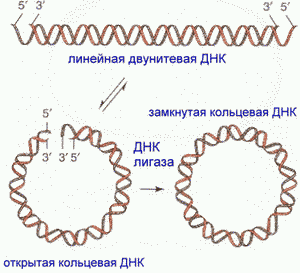
ДНК имеют формы ЛИНЕЙНАЯ, КОЛЬЦЕВАЯ, 2-х и 1-ЦЕПОЧЕЧНАЯ.
Двуцепочечные
ДНК с "липкими" концами могут
образовывать кольцо, которое далее
ковалентно сшивается по сахарофосфатной
цепи при помощи ДНК-лигазы.
Третичная структура ДНК у эукариотических клеток отличается тем, что многократная спирализация ДНК сопровождается образованием комплексов с белками.
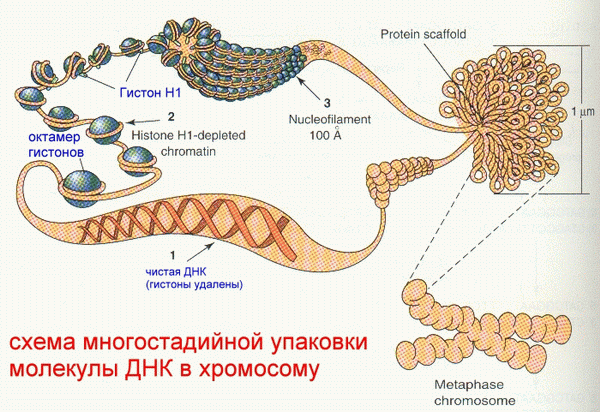
46 хромосом (хроматид) человека организованы в 23 пары. Средняя длина хромосомы составляет 130 млн. пар оснований и имеет длину 5 см. Хромосома №1- 263 млн. п.о., хромосома № 46 –меньше 50 млн. п.о. Если проложить все ДНК в В-конформации в линию, то их общая длина превысит 2 метра. Человеческая хромосома 16 имеет 2,5 мкм в длину, а длина самой ДНК- 3,7 см. Понятно, что уместить такой длины ДНК в ядре возможно только путем ее определенной упаковки. При образовании третичной структуры ДНК человека происходит в среднем уменьшение ее размеров в 100 тысяч раз.
|
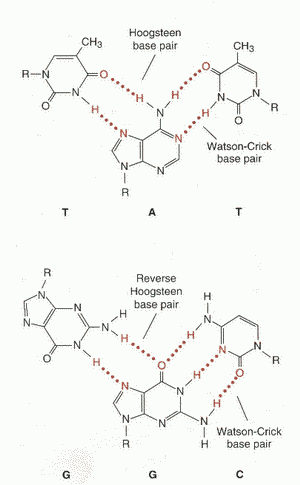
При образовании третичной структуры нуклеиновых кислот возможно образование ТРЕХ- и ЧЕТЫРЕХНИТЕВЫХ участков. Образование ТРЕХНИТЕВЫХ участков благодаря так называемым Хугстэновским взаимодействиям (Hoogsteen base pairs), когда одновременно взаимодействуют три основания: А-А-Т, Т-А-Т, Г-Г-Ц, Ц-Г-Ц. Аналогичным образом происходит образование ТЕТРАМЕРНЫХ участков ДНК. Точный биологический смысл появления трех- и четырехнитевых участков ДНК пока не выяснен. Имеются только лишь предположения о том, что такие участки возникают в местах, наиболее ответственных за процессы репликации и транскрипции. |
|
Материал хромосом- ХРОМАТИН- содержит кроме самой ДНК также гистоны, негистоновые белки, небольшое количество РНК. Нуклеосомный кор содержит октамер гистонов (2 х (Н2а+Н2b+H3+H4)). ГИСТОН- простой белок (примерно 50% хроматина). Нуклеосомный кор образуется при оборачивании октамера гистонов двунитевой спирализованной ДНК на 1,5 оборота, отдельно включается дополнительный белок- гистон Н1. Все вместе называется ХРОМАТОСОМом . Н1- очень богат ЛИЗ, Н2а, Н2b- умеренное количество ЛИЗ, Н3- есть ЦИС, умеренно- АРГ, Н4- богат АРГ и ГЛИ. |
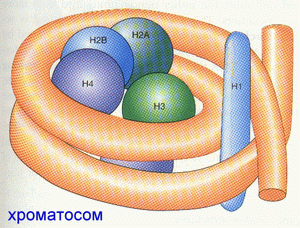
Хроматосомы
образуются на двунитевой спирали ДНК
на дистанциях (называемых ЛИНКЕРами)
от 20 до 90 пар нуклеотидов и напоминают
нанизанные на нитку бусины. Следующий
этап- сворачивание в спираль очень
длинной последовательности “бус”. Эта
спираль, в свою очередь, претерпевает
сворачивание в двужильные канаты, из
которых образуются гроздья, являющиеся
небольшой частью хромосомы: