

Мембранные потенциалы и их ионная природа.
 Опыты
Л. Гальвани и А. Вольта во второй половине
18в. привели
к тому, что функционирование живых
тканей сопровождается электрическими
явлениями. Генерация
и распространение
электрических потенциалов —физическое
явление в живых клетках и тканях.
Биопотенциал-это
разность электрических потенциалов,
образующуюся между двумя точками
клеток, тканей и органов в процессе
их жизнедеятельности. Они отражают
функциональное состояние клеток и
тканей. Их регистрация
и анализ являются важным приемом при
физиологических исследованиях
и в диагностике. Природа
мембранных потенциалов — электрических
потенциалов, образующихся между
внутренней и внешней сторонами мембраны.
Опыты
Л. Гальвани и А. Вольта во второй половине
18в. привели
к тому, что функционирование живых
тканей сопровождается электрическими
явлениями. Генерация
и распространение
электрических потенциалов —физическое
явление в живых клетках и тканях.
Биопотенциал-это
разность электрических потенциалов,
образующуюся между двумя точками
клеток, тканей и органов в процессе
их жизнедеятельности. Они отражают
функциональное состояние клеток и
тканей. Их регистрация
и анализ являются важным приемом при
физиологических исследованиях
и в диагностике. Природа
мембранных потенциалов — электрических
потенциалов, образующихся между
внутренней и внешней сторонами мембраны.
Модельная система, представляющая собой сосуд, разделенный полупроницаемой мембраной . Предположим, в левой части сосуда содержатся ионы калия, хлора и какие-либо крупные частицы, несущие положительный заряд . В правой части сосуда находятся только калий и хлор . Мембрана способна легко пропускать неорганические анионы и катионы, но является непроницаемой для молекул белка. Общее число частиц в растворах одинаково (т.к.сохр. их электронейтральность), однако [K+] втором растворе больше . K+ устремятся из раствора 2 в раствор 1, а вслед за ними пойдут и Cl- (для сохранения электронейтральности растворов). При этом [Cl-]в первом растворе еще больше возрастет. Этот процесс будет продолжаться до установления равновесного состояния (равновесие Доннана). Между двумя сторонами мембраны образуется разность потенциалов, которая уравновешивает концентрационный градиент ионов, способных к диффузии. Мембранная разность потенциалов рассчитывается по формуле Нернста:с1 и с2 — молярные концентрации ионов по обе стороны мембраны, R — универсальная газовая постоянная, Т — термодинамическая температура, при которой происходит диффузия, F — постоянная Фарадея, Z — заряд иона. Эту разность потенциалов называют равновесным мембранным потенциалом.
Мембранная теория происхождения биопотенциалов была выдвинута в 1902 г. Б. Бернштейном. В живой клетке [K+] значительно больше, чем в межклеточной жидкости, и крупные органические молекулы практически не проникают через мембрану. Важным доводом в пользу представлений Бернштейна послужил тот факт, что рассчитанная по формуле Нернста разность потенциалов между наружной и внутренней сторонами мембраны мышечного волокна оказалась близкой к измеренной в опытах с помощью внутриклеточного микроэлектрода. Однако «калиевая теория» мембранного потенциала оказалась несовершенной, не способной объяснить наблюдаемые впоследствии факты отклонения истинных значений потенциалов на мембранах живых клеток от теоретически рассчитанных. Оказалось, что равновесный мембранный потенциал характерен лишь для мертвых клеток, либо клеток с ослабленным метаболизмом. В настоящее время общепризнанной теорией, объясняющей возникновение и поддержание потенциала на клеточной мембране в состоянии физиологического покоя, является теория А. Ходжкина. Она была развита и экспериментально обоснована им в 50-х гг. XX в. Сущность ее заключается в том, что потенциал, существующий на мембранах невозбужденных клеток (потенциал покоя), обусловлен полупроницаемыми свойствами клеточной мембраны и неравномерным распределением ионов между клеткой и окружающей средой. Это распределение поддерживается механизмами активного переноса, локализованными в самой мембране.
Механизм генерации потенциала действия.
В настоящее время общепризнанной теорией, объясняющей возникновение и поддержание потенциала на клеточной мембране в состоянии физиологического покоя, является теория А. Ходжкина. Она была развита и экспериментально обоснована им в 50-х гг. XX в. Сущность ее заключается в том, что потенциал, существующий на мембранах невозбужденных клеток (потенциал покоя), обусловлен полупроницаемыми свойствами клеточной мембраны и неравномерным распределением ионов между клеткой и окружающей средой. Это распределение поддерживается механизмами активного переноса, локализованными в самой мембране.При получении выражения для потенциала покоя важно учитывать знаки ионов, проникающих через мембрану. Это можно сделать, в частности, обозначая знаком плюс плотности потоков положительных ионов и знаком минус — отрицательных. Основной вклад в создание и поддержание потенциала покоя вносят Na+,Cl-.Для живой клетки характерно не равновесное распределение веществ, но существование потоков ионов в обе стороны через мембрану. Cостояние, при котором число различных ионов, проходящих в единицу времени через мембрану внутрь клетки, равно числу выходящих из клетки ионов, называют стационарным. D стационарном состоянии суммарная плотность потока ионов через мембрану равна нулю: J = 0.
![]() уравнение
Гольд мана —
Ходжкина
—
Катца.
уравнение
Гольд мана —
Ходжкина
—
Катца.
Р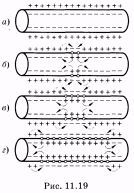 азличные
концентрации ионов внутри и вне клетки
созданы ионными
насосами — системами активного
транспорта. Потенциал
покоя обязан активному переносу.
Потенциал покоя, рассчитанный по формуле
Гольдмана—Ходжкина— Катца, составляет
60 мВ со знаком минус со стороны
внутриклеточного
пространства. На
основании большого экспериментального
материала было установлено, что величина
потенциала покоя может существенно
различаться для разных клеток. Механизм
распространения ПД-основные
положения.
Распространение потенциала действия
вдоль нервного волокна обусловлено
возникновением
так называемых локальных
токов,
образующихся
между возбужденным
и невозбужденным участками
клетки.В
состоянии
покоя (а) внешняя поверхность
клеточной мембраны имеет
положительный потенциал, а внутренняя
— отрицательный. В момент возбуждения
полярность мембраны
меняется на противоположную (б).
В
результате
этого между возбужденным и невозбужденным
участками мембраны возникает разность
потенциалов. Наличие разности потенциалов
и приводит к появлению между этими
участками локальных
токов. На
поверхности клетки локальный ток течет
от невозбужденного
участка к возбужденному; внутри клетки
он течет в обратном
направлении (в). Локальный ток, как и
любой электрический
ток, раздражает соседние невозбужденные
участки и
вызывает увеличение проницаемости
мембраны. Это приводит к возникновению
потенциалов действия в соседних
участках. В то же время
в ранее возбужденном участке происходят
восстановительные
процессы реполяризации. Вновь возбужденный
участок в свою очередь
становится электроотрицательным и
возникающий локальный
ток раздражает следующий за ним участок.
Этот процесс многократно
повторяется и обусловливает распространение
импульсов
возбуждения по всей длине клетки в
обоих направлениях (г).
В
нервной системе импульсы проходят лишь
в определенном направлении из-за
наличия синапсов, обладающих односторонней
проводимостью.
азличные
концентрации ионов внутри и вне клетки
созданы ионными
насосами — системами активного
транспорта. Потенциал
покоя обязан активному переносу.
Потенциал покоя, рассчитанный по формуле
Гольдмана—Ходжкина— Катца, составляет
60 мВ со знаком минус со стороны
внутриклеточного
пространства. На
основании большого экспериментального
материала было установлено, что величина
потенциала покоя может существенно
различаться для разных клеток. Механизм
распространения ПД-основные
положения.
Распространение потенциала действия
вдоль нервного волокна обусловлено
возникновением
так называемых локальных
токов,
образующихся
между возбужденным
и невозбужденным участками
клетки.В
состоянии
покоя (а) внешняя поверхность
клеточной мембраны имеет
положительный потенциал, а внутренняя
— отрицательный. В момент возбуждения
полярность мембраны
меняется на противоположную (б).
В
результате
этого между возбужденным и невозбужденным
участками мембраны возникает разность
потенциалов. Наличие разности потенциалов
и приводит к появлению между этими
участками локальных
токов. На
поверхности клетки локальный ток течет
от невозбужденного
участка к возбужденному; внутри клетки
он течет в обратном
направлении (в). Локальный ток, как и
любой электрический
ток, раздражает соседние невозбужденные
участки и
вызывает увеличение проницаемости
мембраны. Это приводит к возникновению
потенциалов действия в соседних
участках. В то же время
в ранее возбужденном участке происходят
восстановительные
процессы реполяризации. Вновь возбужденный
участок в свою очередь
становится электроотрицательным и
возникающий локальный
ток раздражает следующий за ним участок.
Этот процесс многократно
повторяется и обусловливает распространение
импульсов
возбуждения по всей длине клетки в
обоих направлениях (г).
В
нервной системе импульсы проходят лишь
в определенном направлении из-за
наличия синапсов, обладающих односторонней
проводимостью.
Распространение потенциала действия по миелиновым и безмиелиновым нервным волокнам.
У высокоорганизованных животных с развитой нервной системой толстые волокна оказываются неэкономичными, и затухание сигнала предотвращается другим способом. Мембраны аксонов у них покрыты миелином — веществом, содержащим много холестерина и мало белка . Удельное сопротивление миелина значительно выше удельного сопротивления других биологических мембран.Толщина миелиновой оболочки во много раз больше толщины обычной мембраны, что приводит к возрастанию диаметра волокна и соответственно величины постоянной длины нервного волокна-. Миелиновая оболочка не полностью покрывает все волокно; оно разделено на отдельные сегменты, между которыми на участках длиной около 1 мкм мембрана аксона непосредственно соприкасается с внеклеточным раствором. Области, в которых мембрана контактирует с раствором, называют перехватами Ранвье. В связи с большим сопротивлением миелиновой оболочки по поверхности аксона токи протекать не могут, и затухание сигнала резко уменьшается. При возбуждении одного узла возникают токи между ним и другими узлами. Ток, подошедший к другому узлу, возбуждает его, вызывает появление в этом месте потенциала действия, и процесс распространяется по всему волокну. Затраты энергии на распространение сигнала по волокну, покрытому миелином, значительно меньше, чем по немиелинизированному, так как общее количество ионов натрия, проходящих через мембрану в области узлов, значительно меньше, чем если бы они проходили через всю поверхность мембраны. При некоторых заболеваниях структура миелиновых оболочек нарушается, и это приводит к нарушению проведения нервного возбуждения. При блокировании узлов нервного волокна анестезирующими средствами, сопротивление аксона возрастает и прохождение сигналов по нерву замедляется или совсем прекращается. Поскольку узлы замыкаются через аксоплазму и внеклеточную среду, то можно предположить, что при увеличении сопротивления внешней среды скорость проведения нервного импульса уменьшится. Существует аналогия между распространением ПД по нервному волокну и электромагнитной волной в двухпроводной линии или коаксиальном кабеле. Однако между этими процессами имеется существенное различие. Электромагнитная волна, распространяясь в среде, ослабевает, так как растрачивает свою энергию. Волна возбуждения, проходящая по нервному волокну, не затухает, получая энергию в самой среде (энергию заряженной мембраны). Волны, получающие энергию из среды в процессе распространения, называют автоволнами, а среду — активно-возбудимой средой (ABC).
Энергетические характеристики волны.
При
волновом движении происходит перенос
энергии, которая состоит из кинетической
и потенциальной энергий колеблющихся
частиц среды. Причем потенциальная
энергия обусловлена деформацией
вещества при взаимном смещении частиц.
В отличие от колебаний свободного тела
в волне не происходит взаимного перехода
кинетической и потенциальной энергии
частиц. Мгновенные значения той и другой
энергии изменяются одновременно (в
фазе) соответственно изменению смещения
частиц. Для мгновенного значения энергии
(потенциальной и кинетической) одной
частицы можно записать: ,
,
где
S-
смещение частицы, -
частота колебания частицы, A-
амплитуда колебания частицы, V-
скорость волнового процесса, в котором
участвует частица, m
– масса одной частицы.Из формулы 1.5
следует, что мгновенные значения энергии
каждой частицы среды изменяются во
времени с удвоенной частотой колебания,
причем в каждый момент времени эти
значения для различных частиц отличаются.
Однако среднее значение энергии за
период колебания для всех частиц
одинаково и составляет: eср
= .
.
Рассчитаем энергию волны для некоторого объема V среды, в которой она распространяется.
Если в единице объема среды содержится N частиц, то = Nm —плотность среды и среднее значение энергии волны в объеме V будет:
Еср
=
где
 — объемная плотность энергии
волны.Величина, численно равная средней
энергии Еср,
переносимой волной в единицу времени
t
через заданную поверхность S,
перпендикулярную направлению
распространения волны, называется
потоком
энергии
через эту поверхность:
— объемная плотность энергии
волны.Величина, численно равная средней
энергии Еср,
переносимой волной в единицу времени
t
через заданную поверхность S,
перпендикулярную направлению
распространения волны, называется
потоком
энергии
через эту поверхность:
Ф=

и измеряется в единицах мощности - Вт.Поток энергии, приходящийся на единицу поверхности, называется плотностью потока энергии:

и измеряется в Вт/м2. Плотность потока энергии называют также интенсивностью волны.
В векторной форме:
 .
.
Плотность потока энергии, переносимого волной, можно рассматривать как вектор, совпадающий по направлению с вектором скорости волны.
Вектор
 ,
показывающий направление распространения
волны и равный потоку энергии, проходящему
через единичную площадку, перпендикулярную
этому направлению, называют вектором
Умова:
,
показывающий направление распространения
волны и равный потоку энергии, проходящему
через единичную площадку, перпендикулярную
этому направлению, называют вектором
Умова:
 .
.
Вектор Умова для упругой волны зависит от плотности среды, квадрата амплитуды колебания частиц, квадрата частоты колебаний и скорости распространения волны.
Николай Алексеевич Умов (1846-1915) является исследователем потока энергии. Идеи о движении энергии были изложены в его диссертации "Уравнения движения энергии в телах", защищенной им в 1874 году на физико-математическом факультете Московского университета. И только через десять лет к таким же выводам о движении энергии пришел английский физик Пойнтинг. Имя Умова вошло в историю физики.
Гармонические, затухающие и вынужденные колебания механические колебания.
Колебаниями называются процессы, отличающиеся той или иной степенью повторяемости (качание маятника часов, колебания струны, работа сердца). В зависимости от физической природы повторяющегося процесса различают колебания: механические, электромагнитные, электромеханические. Механические колебания- это колебания, происходящие при отсутствии трения и внешних сил, называются собственными; их частота зависит только от свойств системы.
Гармонические колебания, т.е. такие колебания, при которых колеблющаяся величина (например, отклонение маятника) изменяется со временем по закону синуса или косинуса.
Затухающие колебания, т.е колебания, происходящие в системе при отсутствии внешних сил (но при наличии потерь на трение или излучение), называются свободными. Частота свободных колебаний зависит от свойств системы и интенсивности потерь. Наличие трения приводит к затухающим колебаниям. Колебания с убывающей амплитудой называются затухающими.
Вынужденные колебания – это такие колебания, которые возникают в колебательной системе под действием внешней периодически изменяющейся силы (вынуждающей силы
Резонанс.
Резкого увеличения амплитуды вынужденных колебаний при приближении частоты вынуждающей силы к собственной частоте колеблющегося тела называется резонансом, происходящие при этом колебания - резонансными, а их частота рез — резонансной частотой колебаний.Расчет дает значение резонансной частоты:
рез
=
 .
.
Если очень мало, то p 0 . Подставив рез вместо в (2.13), получим максимальную величину амплитуды колебаний при резонансе:
Арез
=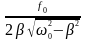 .
.
Чтобы определить резонансную частоту рез, нужно найти максимум функции или минимум выражения, стоящего под корнем в знаменателе. Продифференцировав получим условие, определяющее рез:
-4(02 - 2) + 8 2 = 0.
Это
уравнение имеет три решения:
= 0
и
 .
.
Решение,
равное нулю, соответствует максимуму
знаменателя. Из остальных двух решений
отрицательное должно быть отброшено,
как не имеющее физического смысла
(частота не может быть отрицательной).
Таким образом, для резонансной частоты
получается одно значение:
рез
= .
.
Подставив это значение частоты в (2.13), получим выражение для амплитуды при резонансе:
Арез =
Ультразвук. Методы получения и регистрации.
Ультразвуком называют упругие механические колебания и волны, частота которых превышает 20 кГц, распространяющиеся в форме продольных волн в различных средах. Верхним пределом УЗ частот считают 106 — 107 кГц. Этот предел определяется межмолекулярными расстояниями и поэтому зависит, от агрегатного состояния вещества, в котором распространяется УЗ волна.
Источники и приемники акустических колебаний и ультразвука.
Ультразвук получается с помощью аппаратов, основанных на использовании явлений магнитострикции (при низких частотах) или обратного пьезоэлектрического эффекта (при высоких). Магнитострикция заключается в изменении длины (удлинение и укорочение) ферромагнитного стержня, помещенного в высокочастотное магнитное поле, с частотой изменения направления поля.
Рис. 2.6. Магнитострикционный излучатель УЗ.
1 – волновод, 2 – концентратор звуковой волны, 3 – сердечник, 4 – обмотка магнитострикционного преобразователя, 5 – провода к генератору электрических колебаний.
Обратный пьезоэлектрический эффект заключается в изменении размера (удлинение и укорочение) кристаллической пластинки (кварц, сегнетова соль, титанат бария) под действием высокочастотного электрического поля (до 3 мГц).
Электромагнитные излучатели — получение колебаний подвижной механической системой под действием электромагнита, возбуждаемого переменным током 10200Гц — 12кГц.
Электродинамические излучатели — взаимодействие магнитных полей неподвижного постоянного магнита и звуковой катушки (или стержня), питаемой переменным током (50 — 5000 Гц).
Существуют также и аэро- и гидродинамические излучатели низкочастотного ультразвука.
Приемники УЗ — электроакустические преобразователи. К ним относятся в первую очередь пьезоэлектрические преобразователи, магнитострикционные, полупроводниковые и пьезополупроводниковые, электростатические приемники и электродинамические.
Термические приемники - для измерения интенсивности УЗ.