

3. Конструирование рекомбинантной днк
Сущность генетической инженерии сводится к целенаправленному конструированию генетических систем вне организма с последующим введением их в живой организм. При этом рекомбинантные ДНК становятся составной частью генетического аппарата реципиентного организма и, кроме того, они привносят в него новые генетические и физиолого-биохимические свойства, полезные для человека. К числу таких свойств можно отнести синтез аминокислот и белков, гормонов, ферментов, витаминов и др.

1 Этап сшивание днк
Один из важных этапов конструирования молекулы ДНК — лигирование (или сшивание) генов с помощью фермента ДНК-лигазы. Сшивание фрагментов ДНК, содержащих нужные гены, осуществляют двумя основными методами: а) по «липким» концам; б) с помощью искусственно достроенных «липких» концов.
Сшивание генов (фрагментов) ДНК по «липким» концам, т.е. взаимнокомплементарным участкам, длиной из 4 —6 пар нуклео-тидов, достаточно легко осуществляется ферментом ДНК-лигазой с образованием ковалентной фосфодиэфирной связи между соседними нуклеотидами:

При отсутствии комплементарных «липких» концов у сшиваемых фрагментов их достраивают, т.е. синтезируют искусственно ферментативным путем. Для этой цели применяют так называемые линкеры (или «переходники») — короткие участки ДНК, имеющие разные «липкие» концы:
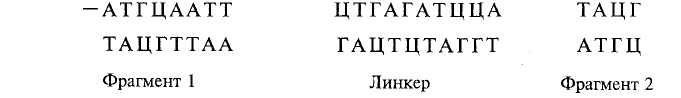
Сшивание генов (фрагментов) ДНК по «тупым» концам
Возможно сшивание фрагментов и по тупым концам, когда концы фрагментов двунитевые:
АТГЦГГГ ЦЦЦГТАЦ
ТАЦГЦЦЦ ГГГЦАТГ
В этом случае реакция лигирования имеет биохимические особенности и ее эффективность ниже, чем при сшивке по «липким» концам.
2. Встраивание в геном
После того как рекомбинантная ДНК сшита, ее вводят в живые клетки. Но поскольку она не способна к самовоспроизведению, ее разрушают внутриклеточные нуклеазы. Для того чтобы рекомбинантная ДНК стала составной частью генетического аппарата клетки, она должна либо встроиться (интегрироваться) в ее геном и реплицироваться за его счет, либо быть способной к автономной репликации. Принято молекулы ДНК, способные акцептировать чужеродную ДНК и автономно реплицироваться, называть векторными молекулами. К числу векторов относят плазмиды, бактериофаги, вирусы животных.
Таким образом, все векторы обеспечивают репликацию встроенных генов, их экспрессию, интеграцию в хромосому клетки и т.д.
Чаще других в генетической инженерии в качестве векторов используют плазмиды.
Плазмидами называют бактериальные репликоны (внехромосомные элементы наследственности), стабильно наследуемые. Они представляют собой двуцепочечные кольцевые молекулы ДНК с вариабельными молекулярными массами. По размеру они соответствуют 1 — 3 % генома бактериальной клетки.

Эукариотические вирусы до сих пор нашли более скромное применение в качестве векторов. Практически используются только онкогенный вирус SV 40 и его производные. Все эти векторы — дефектные вирусы, не способные давать полноценные вирусные частицы в клетке хозяина. Анализируемую ДНК можно вводить и в другие репликоны, способные размножаться в клетках, например бактериофаги.
Векторные плазмиды и векторные вирусы со встроенными чужеродными генами часто называют гибридными (или химерными) плазмидами (или фагами).
1. После конструирования рекомбинантных ДНК их с помощью трансформации вводят в реципиентный организм: бактериальную, грибную, растительную или животную клетку. Трансформация предусматривает предварительную обработку клеток соединениями, обусловливающими проникновение ДНК внутрь клеток с последующим их помещением в среду, в которой способны существовать только клетки, получившие векторную молекулу, например в среду с определенным антибиотиком
2. Процесс инфицирования клеток с помощью чужеродных ДНК, приводящий к образованию зрелого фагового потомства, назван трансфекцией.
Практически общий способ трансформации и трансфекции основан на том, что при обработке клеток бактерий СаС12их мембрана становится проницаемой для ДНК. Однако эффективность проникновения экзогенной ДНК в клетку довольно низка (10%). Поэтому среди бактерий, подвергшихся трансформации, только небольшая часть оказывается трансформированной.
3. Отделение ее от общей массы осуществляется в процессе клонирования. Для клонирования бактериальную суспензию определенной концентрации выливают на твердую питательную среду, например на агар с питательными добавками в чашке Петри из расчета 5 — 10 бактерий на 1 см2 поверхности. Бактериальная клетка на поверхности агара начинает делиться с образованием в итоге маленькой колонии, похожей на шляпку гриба. Эта колония называется клоном, причем из каждой клетки образуется свой клон, все клетки которого имеют свойства бактерии-родоначальника.
Для отделения рекомбинантных бактерий часть материала каждого клона переносят на другую чашку Петри, содержащую антибиотик, ген устойчивости к которому был разрушен при создании рекомбинан-тов. На этих чашках Петри дают клоны только те бактерии, которые содержат исходную плазмиду, а рекомбинантные бактерии их не образуют