

Глава 12. Физиология чувства равновесия, слуха и речи 291
сковых клеток [7, 33, 34], хотя рецепторные потенциалы ими и генерируются, при высокой частоте стимулов регистрируется только постоянное напряжение: мембранный потенциал волосковых клеток не меняется синхронно с высокочастотным звуком. Микрофонный же потенциал:
синхронен звуковому стимулу практически без латентного периода;
лишен рефрактерного периода;
лишен измеримого порога;
не подвержен утомлению; т.е. во всех отношениях отличается от нейронного потенциала действия.
Деполяризация волосковых клеток вызывает выброс из их базальной части медиатора (возможно, глутамата [38]), возбуждающего афферентные нервные волокна. Когда около уха раздается щелчок (короткий импульс давления), волокна слухового нерва активируются синхронно и от круглого окна, помимо микрофонного, можно записать еще и составной потенциал действия. Более длительные звуки вызывают асинхронную импульсацию, не суммирующуюся в отдельные потенциалы действия. На рис. 12.11 показаны микрофонный потенциал улитки (МПУ) и составной потенциал действия (СПД), вызванные щелчком. Они были зарегистрированы на кошках, однако их можно записать и у человека, когда в диагностических целях электрод пропускают через барабанную перепонку и подводят к круглому окну.
Кодирование звука в волокнах слухового нерва.
В улитковом нерве 90% афферентных волокон миелинизированы и отходят от внутренних волосковых клеток. Каждое контактирует только с одной из них, т.е. с очень малой частью улитки. Эти волокна достаточно толсты для регистрации в них с помощью микроэлектродов потенциалов действия и изучения ответа на звуковую стимуляцию (волокна, отходящие от наружных волосковых клеток, для этого слишком тонки). Так как каждый участок улитки соответствует определенной частоте, каждое из таких волокон сильнее всего возбуждается своей характеристической частотой звука, а другими частотами не активируется совсем или активируется лишь при повышении звукового давления. Это отражено на рис. 12.12, где представлен график зависимости порога восприятия от частоты стимула для двух разных волокон. Критерием для установления порога служит определенное повышение активности над ее спонтанным уровнем. Каждое волокно возбуждается при значениях частоты и интенсивности в пределах закрашенной на рисунке области. У ограничивающей ее кривой частотно-пороговой характеристики заметен узкий, заостренный низкопороговый участок и широкий высокопороговый. Частотно-пороговые характеристики отражают распределе-
|
Рис. 12.12. Схематичные частотно-пороговые кривые двух афферентных волокон слухового нерва (а, б) с различными характеристическими частотами (ХЧ). Кривая в типична для волокна с патологическими изменениями, вызванными повреждением внутреннего уха |
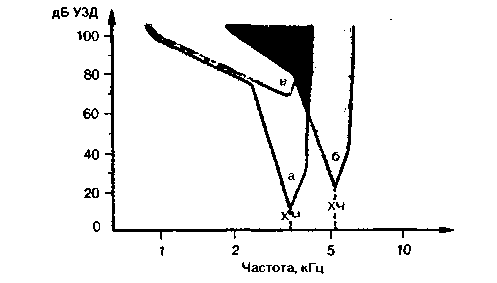
ние частотных максимумов на основной мембране. Реакция одиночного волокна на стимул, выраженная в виде такой кривой, представляет собой спектральный анализ звука. Если в нем несколько различных частот, активируется несколько групп нервных волокон. Длительность звукового стимула кодируется продолжительностью нервной активности, а интенсивность-ее, уровнем. При увеличении звукового давления возрастает и частота импульсации нейронов (до некоторого предела, после чего наступает насыщение). При очень высоком давлении, кроме того, активируются и соседние волокна, находившиеся прежде в состоянии покоя. Такой процесс показан на рис. 12.12; оба волокна возбуждаются, если звук соответствует области перекрывания их частотно-пороговых характеристик. Итак, на уровне первичных афферентов звуковой стимул раскладывается на частотные компоненты. Каждый из них возбуждает соответствующие нервные волокна. На более высоких уровнях слухового тракта нейроны могут вести себя и по-иному.
При повреждениях улитки чувствительность и частотная избирательность афферентных волокон снижаются (рис. 12.12). Рецепторный потенциал внутренних волосковых клеток также меняется и, как говорилось выше, то же происходит с механическими колебательными свойствами основной мембраны. Последние, можно полагать, и определяют поведение этих клеток и волокон, однако сами зависят от процесса активного механического усиления, за который, возможно, отвечают наружные волосковые клетки. Согласно существующей гипотезе, они стимулируются звуком первыми и генерируют дополнительную колебательную энергию той же частоты. Затем она передается внутренним волосковым клеткам. Если это верно, речь идет о своего рода гибриде между сенсорной клеткой и генератором механической энергии [7, 44]. Мно-
292 ЧАСТЬ III. ОБЩАЯ И СПЕЦИАЛЬНАЯ СЕНСОРНАЯ ФИЗИОЛОГИЯ
жество вопросов здесь пока не находит ответа, однако способность улитки как вырабатывать, так и анализировать звуковую энергию бесспорна. Звук, генерируемый в улитке, можно измерить даже снаружи от барабанной перепонки [7, 36]. Эти процессы очень часто нарушаются при различных повреждениях (см. с. 295).
Наружные волосковые клетки подходят на роль активных усилителей, поскольку содержат сократительные белки и обслуживаются чрезвычайно развитой эфферентной сетью. Кроме того, их афференты явно не существенны для передачи информации в мозг [7, 13].
Кодирование звуковых частот по принципу локализации рецептора рассмотрено выше. Второй вид кодирования информации в слуховом нерве следующий. Тоны частотой до 5 кГц обычно вызывают нейронную импульсацию в слуховом нерве только в определенные фазы цикла звукового колебания. В результате временная структура стимула (например, период Τ на рис. 12.5, Б) представлена группами потенциалов действия, передаваемых в ЦНС по слуховому нерву в соответствующие моменты времени. Мозг, очевидно, способен оценивать временную структуру импульсации и определять лежащую в его основе звуковую частоту (так называемый анализ периодичности). Особенно четкие доказательства этого были получены при прямой электростимуляции слухового нерва пациентов, страдающих глухотой: периодические стимулы перерабатывались таким образом, что возникало ощущение тона определенной высоты, что показывает действительную важность анализа периодичности для слуха [30, 46].