

2.3. Нервная система позвоночных
У всех позвоночных животных общий принцип построения нервной системы резко отличен от узлового типа построения нервной системы беспозвоночных животных. Центральная нервная система позвоночных животных с самых ранних этапов эмбрионального развития не имеет узлового строения; она закладывается в виде сплошной нервной трубки, которая затем дифференцируется на различные отделы мозга и которая в дальнейшем является также источником образования нервных узлов на периферии (в симпатической и парасимпатической нервных системах). Такие узлы являются, следовательно, вторичными образованиями и состоят из клеток, мигрировавших из первоначальной сплошной нервной трубки. Нервная трубка всегда расположена на спинной (дорсальной) стороне тела, в то время как узловая нервная цепочка сложных беспозвоночных является брюшной и расположена на вентральной стороне.
У наиболее примитивных форм хордовых (ланцетник) центральная нервная система сохраняет характер мало дифференцированной сплошной нервной трубки и во взрослом состоянии. У всех же более сложно организованных форм происходит дифференциация последней. Она теряет свою равномерную форму, и на ее головном конце возникают утолщения нервной ткани в виде пузырей. Эти пузыри являются эмбриональным зачатком головного мозга.
Рассматривая различные классы позвоночных и сравнивая у них типы строения центральной нервной системы, довольно легко убедиться в том, что у всех образование мозговых пузырей происходит по одному и тому же типу; можно проследить эволюцию каждого пузыря, начиная от самых примитивных форм позвоночных и кончая млекопитающими. Варьирует лишь степень его развития и степень дифференциации в нем нервных элементов.
Основой образования головного мозга является деление переднего отдела мозговой трубки на 3 части, дающих начало трем мозговым пузырям, которые отчетливо выделяются у самых примитивных из существующих позвоночных — у круглоротых (следует учесть, что у современных круглоротых, особенно у миксин, есть черты деградации в связи с полупаразитическим образом жизни). По расположению эти эмбриональные пузыри можно обозначить как передний, средний и задний; соответственно у взрослого животного они образуют передний, средний и задний мозг.
У круглоротых задний и средний пузыри уже достаточно хорошо развиты и имеют весьма сложную нейронную структуру. Задний пузырь у миноги образует продолговатый мозг и, хотя и небольшой по объему, но хорошо видимый мозжечок, который имеет форму поперечного валика. Средний пузырь дифференцируется в весьма сложный средний мозг, который еще не разделяется на отдельные холмы, как у более высокоорганизованных животных. Он является основным центром обработки зрительных сигналов. Передний же мозговой пузырь сохраняет малодифференцированный характер. Из него образуются обонятельные луковицы и обонятельные доли, в которых заканчиваются пути от рецепторов обонятельной ямки; они тесно прижаты к слаборазвитому промежуточному мозгу.
Для рыб характерен примерно такой же общий план строения головного мозга — сложно организованный задний и средний мозг и мало дифференцированный (хотя и больший по размерам) передний мозг, выполняющий обонятельную функцию. В связи с тем, что движения у рыб значительно более сложны, и многим из них для существования необходимы точное поддержание положения тела в воде и тонкая двигательная координация, у них очень интенсивно развиваются те участки заднего мозга, которые связаны с высшей координацией двигательных актов, в частности — мозжечок. Мозжечок у многих рыб становится доминирующей частью центральной нервной системы (рис. 1.4).
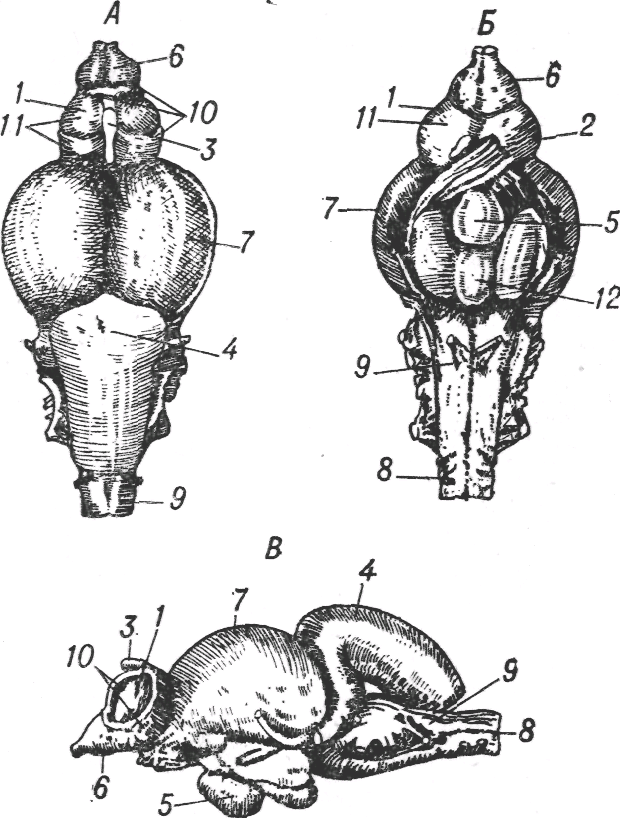
Рис. 1.4. Мозг форели. Вид сверху (А), снизу (Б) и сбоку (В);1 — базальный ганглий, 2 — хиазма, 3 — эпифиз, 4 — мозжечок, 5 — гипофиз, 6 — обонятельные доли, 7 — средний мозг, 8 — спинной мозг, 9 — задний мозг, 10 — плащ переднего мозга, 11 — передний мозг.
После выхода позвоночных на сушу происходит перераспределение функционального значения различных отделов головного мозга и соответственно изменение объема и сложности образований, возникающих из каждого мозгового пузыря. Это можно хорошо видеть, сравнивая мозг амфибий и рептилий с мозгом рыб. Задний мозг у первых занимает относительно меньший объем; мозжечок также развит значительно меньше, зато средний мозг развивается очень значительно, усложняясь в структурном отношении. Наряду с развивающимся в нем двухолмием, являющимся высшим зрительным центром, появляются дополнительные бугорки, в которые направляются пути от развившихся у этих животных органов слуха. Сравнительно простое строение мозжечка у амфибий может быть связано с упрощением этих животных, класс амфибий на данном этапе эволюционного развития находится на стадии деградации, ныне живущие земноводные отличаются весьма простыми, даже примитивными движениями.
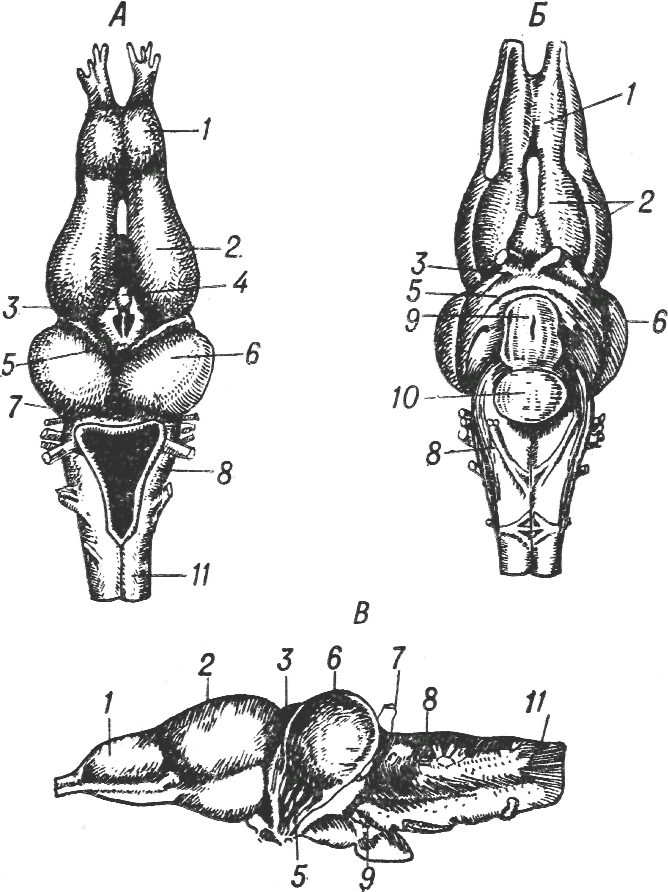
Рис. 1.5. Мозг лягушки (по Р. Видерсгейму)- Вид сверху (А), снизу (Б) и сбоку (В); 1 — обонятельные доли, 2 — передний мозг, 3 — промежуточный мозг, 4 — эпифиз, 5 — зрительный тракт, 6 — средний мозг, 7 — мозжечок, 8 — задний мозг, 9 — воронка, 10 — гипофиз, 11 — спинной мозг.
И у амфибий, и у рептилий резко увеличивается передний мозг, который приобретает сложную анатомическую форму и не менее сложную внутреннюю структуру. Передний мозг дифференцируется на промежуточный мозг и два симметричных полушария, с которыми связаны обонятельные луковицы (рис. 1.5). Хотя передний мозг оказывается все еще осуществляющим одну из рецепторных функций, а именно — обоняние, он приобретает и более сложные координирующие функции, которые у рыб целиком связаны со средним мозгом.
С дальнейшим эволюционным прогрессом позвоночных животных связаны две различные линии развития мозговых пузырей.
Одна линия — это линия млекопитающих, у которых развитие нервной системы идет в сторону чрезвычайного увеличения и усложнения наружной части переднего мозгового пузыря, т. е. формирующей кору больших полушарий (рис. 1.6). Наряду с этим интенсивно развиваются и связанные с корой больших полушарий новые отделы мозжечка, который становится довольно объемистым, уступающим по массе нервной ткани и по сложности ее организации только структурам переднего мозгового пузыря. Остальные мозговые пузыри, в частности средний, явно отстают в развитии. Основные пути передачи зрительных сигналов, которые раньше направлялись в средний мозг, теперь через промежуточный мозг отклоняются в кору больших полушарий.
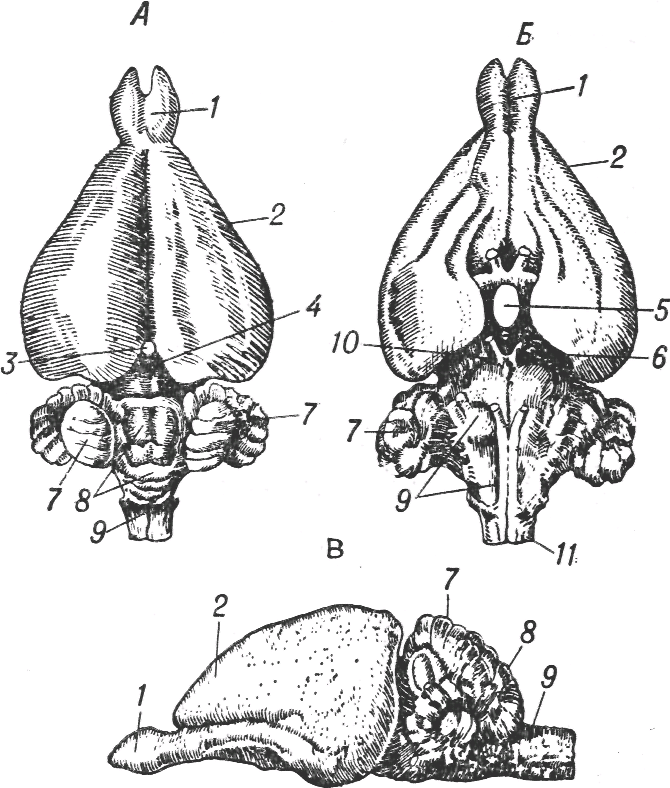
Рис. 1.6. Мозг кролика (по Р. Видерсгейму). Вид сверху (А), снизу (Б) и сбоку (В); 1 — обонятельные луковицы, 2 — передний мозг, 3 — эпифиз, 4 — средний мозг, 5 — гипофиз, 6 — хиазма, 7 — полушария мозжечка, 8 — червь мозжечка, 9 — задний мозг, 10 — ножки мозга, 11; — спинной мозг.
Вторая линия развития центральной нервной системы представлена головным мозгом птиц. Для этих животных характерно умеренное развитие из переднего мозгового пузыря коры больших полушарий; доминирующими по сложности и по объему остаются структуры, которые спрятаны в глубине полушарий (базальные ганглии и промежуточный мозг).