

Визначення стартових та кінцевих точок і рівня активності транскрипції
Кілька описаних нижче прикладів ілюструють деякі підходи, що розроблено для характеристики транскриптів та дослідження рівня транскрипційної активності.
Картування за допомогою нуклеази S1 дозволяє не тільки визначати старт та кінцеву точку транскрипції певного гена, а й оцінювати рівень транскрипційної активності цього гена. Процес починається з клонованого фрагмента гена, який містить кінцеву точку, що має бути встановленою. Навкруг неї обирають два рестриктні сайти однієї рестриктази, і ще один – іншої, як показано на рис. 11.12. Перша рестриктаза вирізає фрагмент, залишаючи одноланцюгові вирости на 5'-кінцях. Використовуючи їх у якості матриці, фрагмент Кленова ДНК-полімерази І добудовує 3'-кінці, приєднуючи радіоактивно-мічені нуклеотиди – у результаті 3'-кінці обох ланцюгів містять мітку. Щоб позбавитись однієї з них, використовують сайт другої рестриктази, який все ще залишається всередині. Нарешті маємо мітку на 3'-кінці тільки матричного ланцюга (який з ланцюгів є матричним, треба знати при підборі рестриктаз або встановити по кінцевому результату). Денатурація цього фрагменту дає радіоактивно-мічений одноланцюговий зонд, для якого є точно відомими його довжина та локалізація у гені. На останніх етапах проводять гібридизацію цього зонда із сумарним препаратом клітинної РНК у розчині – комплементарна частина відповідної мРНК утворює із зондом подвійну спіраль. Після цього й використовують нуклеазу S1, яка здатна гідролізувати тільки одноланцюгову нуклеїнову кислоту. Для локалізації кінцевої точки транскрипції залишається визначити точну довжину отриманого міченого фрагмента ДНК за допомогою електрофорезу: ця довжина дорівнює відстані від мітки до кінцевої точки. Інтенсивність смуги після електрофорезу дає інформацію про транскрипційну активність гена: чим вищою є інтенсивність, тим більше мРНК містилося у препараті, виділеному з клітини.
 Рис.
11.12. S1-картування
точки закінчення транскрипції.
Рис.
11.12. S1-картування
точки закінчення транскрипції.
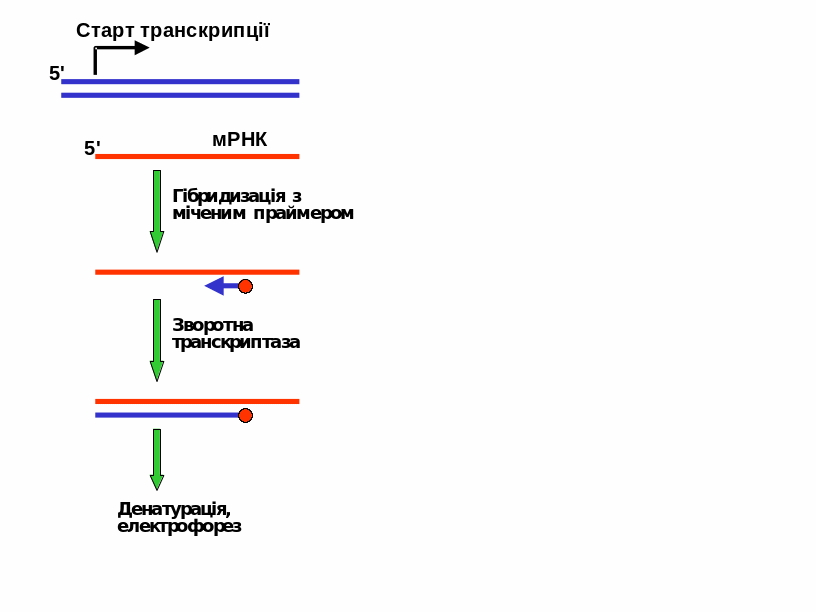 Рис.
11.13. Метод
подовження праймера для визначення
старту транскрипції.
Рис.
11.13. Метод
подовження праймера для визначення
старту транскрипції.
Визначення транскрипційної активності in vivo. Описані вище методи визначають рівень присутності мРНК певного типу в клітині. Цей рівень залежить, насправді, не тільки від рівня транскрипційної активності, але й від швидкості деградації мРНК. Один з методів, що дозволяє встановити, чи відбувається транскрипція в даний момент, полягає у тому, що клітинні ядра виділяються і до них додаються мічені нуклеозидтрифосфати. Нові акти ініціації вже не відбуваються у таких відокремлених ядрах, і РНК-полімераза просто продовжує добудовувати транскрипти, синтез яких розпочався до виділення. Зрозуміло, що такі подовжені транскрипти виявляються міченими. Після виділення РНК їх треба ідентифікувати, що можна зробити шляхом гібридизації з ДНК генів, які цікавлять дослідника.