

Курс лекций "Продуктивность экосистем"
Энергия в экосистемах
Напомним, что экосистема - это совокупность живых организмов, обменивающихся непрерывно энергией, веществом и информацией друг с другом и с окружающей средой.
Энергию определяют как способность производить работу. Свойства энергии описываются законами термодинамики.
Первый закон (начало) термодинамики или закон сохранения энергии утверждает, что энергия может переходить из одной формы в другую, но она не исчезает и не создается заново.
Второй закон (начало) термодинамики или закон энтропии утверждает, что в замкнутой системе энтропия может только возрастать. Применительно к энергии в экосистемах удобна следующая формулировка: процессы, связанные с превращениями энергии, могут происходить самопроизвольно только при условии, что энергия переходит из концентрированной формы в рассеянную, то есть деградирует. Мера количества энергии, которая становится недоступной для использования, или иначе мера изменения упорядоченности, которая происходит при деградации энергии, есть энтропия. Чем выше упорядоченность системы, тем меньше ее энтропия.
Самопроизвольные процессы ведут систему к состоянию равновесия с окружающей средой, к росту энтропии, производству положительной энтропии. Если неживую неуравновешенную с окружающей средой систему изолировать, то всякое движение в ней скоро прекратится, система в целом угаснет и превратится в инертную группу материи, находящуюся в термодинамическом равновесии с окружающей средой, то есть в состоянии с максимальной энтропией. Это наиболее вероятное для системы состояние и самопроизвольно без внешних воздействий она выйти из него не сможет. Так, например, раскаленная сковородка, остыв, рассеяв тепло, сама уже не нагреется; энергия при этом не потерялась, она нагрела воздух, но изменилось качество энергии, она уже не может совершать работу. Таким образом, в неживых системах устойчиво их равновесное состояние.
У живых систем есть одно принципиальное отличие от неживых - они совершают постоянную работу против уравновешивания с окружающей средой. Это утверждение имеет следующий термодинамический смысл: в живых системах устойчиво неравновесное состояние.
Жизнь - это единственный на Земле естественный самопроизвольный процесс, в котором энтропия системы уменьшается. Почему это возможно?
Все живые системы являются открытыми для обмена энергией. В окружающей их среде есть огромное количество даровой энергии Солнца, а в составе самой живой системы есть компоненты, обладающие механизмами, позволяющими эту энергию улавливать (извлекать), концентрировать, а затем снова рассеивать в окружающую среду. Рассеивание энергии, то есть увеличение энтропии, - это процесс, характерный для любой системы, как неживой, так и живой, а самостоятельное улавливание и концентрирование энергии - это способность только живой системы. При этом происходит извлечение порядка, организации из окружающей среды, то есть выработка отрицательной энергии - негоэнтропии. Такой процесс образования порядка в системе из хаоса окружающей среды называется самоорганизацией. Он ведет к уменьшению энтропии живой системы, противодействует ее уравновешиванию с окружающей средой, то есть росту энтропии, что для живой системы при достижении максимальной энтропии - равновесия с окружающей средой - означает смерть.
Таким образом, любая живая система, в том числе и экосистема, поддерживает свою жизнедеятельность благодаря, во-первых, наличию в окружающей среде в избытке даровой энергии; во-вторых, способности за счет устройства составляющих ее компонентов эту энергию улавливать и концентрировать, а использовав - рассеивать в окружающую среду.
Даровая энергия окружающей среды - это энергия Солнца. Доходящая до Земли энергия Солнца распределяется следующим образом: 33 % ее отражается облаками и пылью атмосферы (это так называемое альбедо или коэффициент отражения Земли); 67 % поглощается атмосферой, поверхностью Земли и океаном. Из этого количества поглощенной энергии лишь около одного процента расходуется на фотосинтез, а вся остальная энергия, нагрев атмосферу, сушу и океан, переизлучается в космическое пространство в форме невидимого теплового (инфракрасного) излучения. Этого одного процента энергии достаточно для обеспечения ей всего живого вещества планеты и поддержания им состояния с низкой энтропией. Как распределяется эта энергия между компонентами биотической структуры?
Улавливают энергию Солнца и превращают ее в потенциальную энергию органического вещества растения - продуценты. Весь остальной живой мир получает необходимую для жизнедеятельности энергию, в основном поедая их.
Перенос энергии пищи от ее источника - продуцента через ряд организмов, происходящий путем поедания одних организмов другими, называется пищевой или трофической цепью.
Содержащееся в пище органическое вещество расщепляется в присутствии кислорода с выделением энергии. Этот процесс, обратный фотосинтезу, называется дыханием. Он имеет место в каждой клетке живого организма, поэтому его еще называют клеточным дыханием.
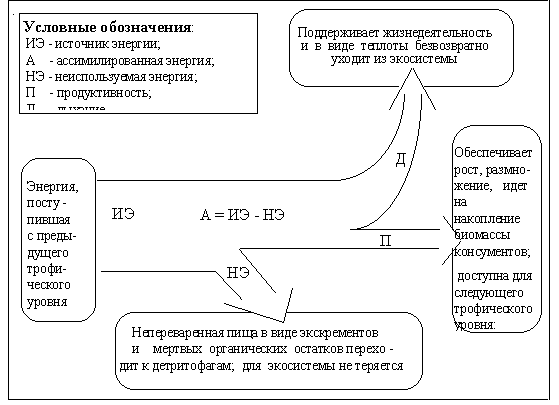
Около 90 % выделившейся энергии расходуется организмом на поддержание своей жизнедеятельности, то есть на обеспечение всех необходимых ему функций, после чего она в виде выделяемого организмом тепла рассеивается в окружающую среду и по сути дела безвозвратно теряется для всей живой системы. И только около 10 % энергии идет на построение тела, рост и размножение организма. Именно эти 10 % энергии и доступны следующему трофическому уровню. Таким образом, энергии с переходом от одного уровня к другому остается все меньше.
Но здесь нужно иметь в виду, что чем выше трофический уровень, тем в более концентрированной форме содержится в живых организмах энергия. Это объясняется присущей только живому веществу спецификой - обладанием механизмами концентрирования энергии.
Таким образом, сначала улавливание, а затем концентрирование энергии с переходом от одного трофического уровня к другому обеспечивает повышение упорядоченности, организации живой системы, то есть уменьшение ее энтропии. Для поддержания низкой энтропии в равной степени важно, чтобы у элементов системы были эффективные механизмы как для улавливания и концентрации энергии - извлечения негоэнтропии из окружающей среды, так и для рассеивания ее в окружающую среду - освобождение от накапливающейся положительной энтропии. В таком сочетании они есть только в живых системах. Поэтому жизнь как термодинамический процесс представляет собой непрерывный обмен живых систем с окружающей средой, при котором происходит освобождение от производимой положительной энтропии и извлечение отрицательной, то есть порядка и организации.
Необходимо понимать, что энтропия уменьшается в конкретной локальной зоне, при этом в окружающей среде она возрастает. Таким образом, рост упорядоченности в одной части системы приводит к усилению неупорядоченности в других ее частях.
Для описания поведения энергии в экосистемах употребляют термин поток энергии, поскольку в отличие от циклического движения веществ превращения энергии идут в одном направлении. Энергия, однажды использованная каким-либо организмом, превращается в тепло и утрачивается для экосистемы. Для своей жизнедеятельности каждый живой компонент, будь то организм или экосистема, должен получать от окружающей среды на входе постоянный приток дополнительной энергии. Живые замкнутые термодинамические системы невозможны.
Энергия и продуктивность экосистем
Итак, жизнь в экосистеме поддерживается благодаря непрекращающемуся прохождению через живое вещество энергии, передаваемой от одного трофического уровня к другому; при этом происходит постоянное превращение энергии из одних форм в другие. Кроме того, при превращениях энергии часть ее теряется в виде тепла.
Весь запас энергии сосредоточен в массе органического вещества - биомассе, поэтому интенсивность образования и разрушения органического вещества на каждом из уровней определяется прохождением энергии через экосистему (биомассу всегда можно выразить в единицах энергии).
Скорость образования органического вещества называют продуктивностью. Различают первичную и вторичную продуктивность.
Энергия поступает в живую составляющую экосистемы через продуценты. Скорость накопления энергии продуцентами в форме органического вещества, которое может быть использовано в пищу, называется первичной продуктивностью. Этим показателем определяется общий поток энергии через живую составляющую экосистемы, а значит, и количество (биомасса) живых организмов, которые могут существовать за ее счет в экосистеме.
В первичной продуктивности различают валовую и чистую продуктивность. Валовая первичная продуктивность - это скорость, с которой растения накапливают химическую энергию при фотосинтезе. Часть ее - около 20 % - они тратят на дыхание - поддержание собственной жизнедеятельности, которая затем в виде теплоты выделяется в окружающую среду и теряется для экосистемы. Скорость накопления органического вещества продуцентами за вычетом расхода на дыхание называется чистой первичной продуктивностью. Это энергия, которую могут использовать организмы следующих трофических уровней.
Поступившая на уровень консументов любого уровня энергия распределяется следующим образом (рис. 1).
Скорость накопления органического вещества на уровнях консументов называется вторичной продуктивностью. Это энергия, которую могут использовать консументы следующего трофического уровня.
Из рассмотренного механизма передачи энергии по цепи живого вещества в экосистеме видно, что в каждом звене пищевой цепи часть энергии - около 90 % - теряется. Поэтому длина пищевой цепи ограничивается размерами этих потерь и, как правило, составляет 3 - 4 уровня.
При этом с повышением трофического уровня его биомасса снижается, так как, во-первых, если фитофаги будут потреблять больше биомассы, чем ее производят продуценты, что, например, имеет место при избыточном выпасе, то популяция продуцентов в конце концов исчезнет; во-вторых, существенная доля потребляемой консументами биомассы не усваивается и возвращается в экосистему в виде экскрементов, а из той, что усваивается, лишь несколько процентов идет на создание биомассы. Таким образом, в естественных экосистемах на высших трофических уровнях не может быть большой биомассы.
В результате последовательности превращений энергии в пищевых цепях каждое сообщество живых организмов в экосистеме приобретает определенную трофическую структуру. Трофическая структура сообщества отражает соотношение между продуцентами, консументами (отдельно первого, второго и т.д. порядков) и редуцентами, выраженное или количеством особей живых организмов, или их биомассой, или заключенной в них энергией, рассчитанных на единицу площади в единицу времени. Графически трофическую структуру сообщества представляют в виде пирамиды. Основанием пирамиды служит первый трофический уровень - уровень продуцентов, а последующие уровни образуют следующие этажи пирамиды. При этом высота всех блоков-этажей - одинакова, а длина пропорциональна числу, биомассе или энергии на соответствующем уровне (рис.).
В зависимости от того, количественное соотношение каких величин отражает пирамида, она называется пирамидой чисел, биомасс или энергий.
Пирамиды чисел и биомасс отражают статику системы, то есть характеризуют количество или биомассу организмов в определенный промежуток времени. Они не дают полной информации о трофической структуре экосистемы, хотя также позволяют решать ряд практических задач, особенно связанных с сохранением устойчивости экосистем. Пирамида чисел позволяет, например, рассчитывать допустимую численность отстрела животных в охотничий период без последствий для нормального функционирования экосистемы.
Из количественных оценок, связанных с энергией, для трофических цепей известно правило десяти процентов: с одного трофического уровня экологической пирамиды энергий на другой в среднем переходит около 10 % энергии, поступающей на предыдущий уровень.
Пирамида энергий является наилучшим графическим изображением трофической структуры экосистемы, поскольку она отражает динамику системы, то есть скорость прохождения энергии через пищевую цепь. Представление о трофической структуре экосистем позволяет выяснить, чем определяется реальная продуктивность экосистемы.
В любой экосистеме происходит образование биомассы и ее разрушение, причем эти процессы всецело определяются жизнью низшего трофического уровня - продуцентами. Все остальные организмы только потребляют уже созданное растениями органическое вещество и, следовательно, общая продуктивность экосистемы от них не зависит.
В зеленых тканях растений осуществляется два параллельных, но противоположных процесса - фотосинтез и дыхание. При фотосинтезе вещество создается, энергия накапливается, а при дыхании часть накопленных веществ и энергии расходуется. Поэтому дыхание рассматривается как некоторая мера энергии, выносимой из сообщества, в то время как увеличение биомассы и есть продуктивность.
Теперь понятно, что если в экосистеме процессы накопления вещества преобладают над процессами дыхания, то есть отношение продуцируемой массы Р к расходам на дыхание Д больше единицы (Р/Д > 1), то суммарная биомасса в ней нарастает. Если же в процессе дыхания или потребления последующими звеньями пищевой цепи расходуется больше вещества, чем создается растениями (Р/Д < 1), то запасы биомассы убывают. При равенстве биомассы, продуцируемой растениями в процессе фотосинтеза и убывающей за счет дыхания тех же растений или расходования последующими уровнями (Р/Д=1), объем биомассы в ней остается примерно постоянным.
Высокие скорости продуцирования биомассы наблюдаются в естественных и искусственных экосистемах там, где благоприятны абиотические факторы, и особенно при поступлении дополнительной энергии извне, что уменьшает собственные затраты системы на поддержание жизнедеятельности. Такая дополнительная энергия может поступать в разной форме: например, на возделываемом поле - в форме энергии ископаемого топлива и работы, совершаемой человеком или животным.
Оценивая продуктивность экосистемы, зависящую от соотношения Р/Д, необходимо учитывать как утечки энергии, связанные со сбором урожая, загрязнением среды, неблагоприятными климатическими условиями и с другими типами стрессовых воздействий, способствующих отведению энергии от процесса продукции - увеличению Д, так и поступления энергии, которые увеличивают продуктивность Р, компенсируя потери тепла при дыхании - при "откачивании неупорядоченности", необходимом для поддержания биологической структуры.
Таким образом, для обеспечения энергией всех особей сообщества живых организмов экосистемы необходимо определенное количественное соотношение между продуцентами, консументами разных порядков, детритофагами и редуцентами. Однако для жизнедеятельности любых организмов, а значит и системы в целом, только энергии недостаточно, они обязательно должны получать различные минеральные компоненты, микроэлементы, органические вещества, необходимые для построения молекул живого вещества.
Высокие скорости продуцирования встречаются в естественных и искусственных экосистемах там, где физические факторы благоприятны, и особенно при дополнительном поступлении энергии извне, уменьшающем собственные затраты на поддержание жизнедеятельности. Такие поступления энергии могут иметь место в разной форме: в тропическом дождевом лесу — в форме работы ветра и дождя, в эстуарии — в форме энергии прилива, на возделываемом поле - в форме энергии ископаемого топлива и работы, совершаемой человеком или животными.
Оценивая продуктивность экосистемы, необходимо учитывать как утечки энергии, связанные со сбором урожая, загрязнением среды, неблагоприятными климатическими условиями и другими типами стрессовых воздействий, способствующих отведению энергии от экосистемы, так и поступления энергии, которые увеличивают продуктивность, компенсируя потери тепла при дыхании (при «откачивании неупорядоченности»), необходимые для поддержания биологической структуры.
Оценки продуктивности особенно важны для сельского хозяйства. На примере возделывания сои можно посмотреть, каким образом распределяется первичная валовая продукция урожая сои. Около 25 % расходуется на дыхание, 5% потребляются симбиотическими микроорганизмами, 5 % потребляют насекомые-вредители (с учетом использования пестицидов), 32 % выносится из данной экосистемы человеком (собственно урожай бобов сои), оставшиеся 33 % в стеблях, листьях и корнях разлагаются в почве и подстилке. По отношению к экосистеме урожай, собранный человеком, является утечкой энергии, обедняющей экосистему.
Столь высокое отношение чистого урожая к валовому продукту достигается только за счет дополнительных вложений энергии, затрачиваемой на обработку земли, орошение, удобрение, селекцию, борьбу с вредителями и т.п. Всякое дополнительное вложение энергии, увеличивающее продуктивность экосистемы, называется энергетической субсидией.
Энергетические субсидии не обязательно организуются человеком. Самый простой пример природной энергетической субсидии является ветер. Ранее уже говорилось, что на приведение в движение круговорота воды тратится около одной трети солнечной энергии, поступающей на Землю. Неравномерный прогрев поверхности планеты приводит к перемещению воздушных масс, что обеспечивает перенос влаги с океана на континенты. Это один из путей дополнительной откачки энергии из солнечного света помимо фотосинтеза. Аналогичные энергетические субсидии может организовать и человек путем искусственного орошения.
Природные энергетические субсидии от энергии приливов получают прибрежные зоны океана. Приливы обеспечивают перенос минеральных веществ, пищи и отходов. В результате некоторые прибрежные зоны характеризуются такой же валовой продуктивностью, как и обрабатываемые человеком поля.
Возможность получать высокие урожаи, совершенно немыслимые еще 100 лет назад, человек обеспечил только за счет энергетических субсидий в искусственно созданные им экосистемы (агроценозы). При этом он селекционировал новые высокоурожайные сорта сельскохозяйственных культур, выращивание которых оправдано только при наличии таких субсидий. Это, кстати, является причиной некоторых неудач при попытках возделывать такие сорта в бедных странах. В США, например, не каждую калорию полученной пищи вкладывается примерно 10 калорий энергии топлива. Для удвоения урожая дополнительные поступления энергии необходимо увеличить где-то в 10 раз. Могут ли такое себе позволить бедные страны, где до сих пор в сельском хозяйстве преобладает немеханизированный труд?
Нужно отметить, что чрезмерные энергетические субсидии могут понизить продуктивность экосистемы. Например, распашка земель в северных широтах является необходимым компонентом сельского хозяйства, в тропических зонах это приводит к быстрому выщелачиванию почвы и потере в ней органического вещества. Проточная вода может способствовать росту продуктивности экосистемы ручья по сравнению с экосистемой в стоячих водах, но слишком быстрое течение может оказаться разрушительным.
Оценивая ситуацию в мире, надо сказать, что ввиду отсутствия достаточных взвешенных энергетических субсидий в развивающихся странах урожаи растут медленнее, чем численность населения. В результате с каждым годом растет число стран, которые вынуждены ввозить продукты питания. Таким образом, без энергетических субсидий в производство пищи человеческая цивилизация существовать уже не в состоянии. И с каждым годом эта ситуация будет усугубляться. При этом где-то впереди опять маячит призрак тепловой катастрофы, избежать которой нам, по-видимому, вряд ли удастся.
Все искусственные экосистемы характеризуются тем, что определенное количество продукции изымается из экосистемы человеком. Если не возмещать эти потери в форме энергетических субсидий, то рано или поздно экосистема деградирует. Например, на полях структура почвы может быть разрушена настолько, что потребуются сотни лет, чтобы на этих землях без участия человека заново возродилась нормальная жизнь. В отличие от агроценозов в естественных экосистемах существует равновесие между производством биомассы и ее разложением. Продукты разложения вновь поступают в круговорот, обеспечивая длительное существование данной экосистемы (такая экосистема, как биогеоценоз, считается теоретически бессмертной).
Биомасса и продуктивность экосистем определят не только количество урожая, которое мы можем снять с данной экосистемы. От этих показателей зависит средообразующая и средостабилизирующая роль экосистем. Например, от этих параметров экосистем напрямую зависит углекислотный баланс в атмосфере. Так при образовании 1т растительной продукции поглощается 1,5-1,8 т углекислого газа с выделением 1,2-1,4 т кислорода. От этих параметров зависит также пылеосаждающая способность леса. Суммарная поверхность листьев в лесу в 10-15 раз превышает занимаемую им площадь, что позволяет осенью осаждать на землю до 50-60 т пыли с гектара. Концентрационная функция живого вещества позволяет очищать атмосферу от вредных химических агентов. Кроме того, с объемом биомассы связана водорегулирующая функция леса и т.п.
ПЕРВИЧНАЯ ПРОДУКЦИЯ
Первичная продукция лежит в основе всей трофической структуры сообщества. Все механизмы экосистемы приводятся в движение энергией, поставляемой фотосинтезом. Поток энергии через популяции растительноядных, плотоядных и детритоядных животных и происходящие в экосистеме круговороты питательных веществ в конечном счете зависят от первичной продуктивности растений.
В состав растений входит не одна глюкоза (что следует из известного уравнения фотосинтеза). В различных биохимических процессах глюкоза используется не только в качестве источника энергии, но и в качестве материала для построения других более сложных органических соединений. В результате соответствующих перестроек сахара превращаются в жиры, масла и целлюлозу - основной материал, из которого состоят стенки растительных клеток. Азот, фосфор, сера и магний, соединяясь с углеродом, кислородом и водородом, образуют целый ряд белков, нуклеиновых кислот и пигментов.
Хлорофилл, помимо атомов азота и углерода, содержит атом магния, подобно тому, как гемоглобин содержит атом железа. Таким образом, даже если все другие необходимые материалы имелись бы в достатке, растение, лишенное магния, не могло бы продуцировать хлорофилл, а тем самым не могло бы расти. Совершенно ясно, что растения не могут также функционировать и в отсутствие воды. Количество воды, необходимое для фотосинтеза, - это всего лишь капля по сравнению с тем огромным ее количеством, которое необходимо для того, чтобы покрыть расходы на транспирацию. Ткани растений состоят главным образом из воды, и вода служит той средой, в которой должны быть растворены питательные вещества, для того чтобы они стали доступными растениям.
В основное уравнение продукции должно входить не только сырье, необходимое для фотосинтеза, но и ряд минеральных веществ. В создании продукции участвует много биологических реакций, но в общем виде этот процесс можно свести к следующему суммарному уравнению:
СО2 + Н2О + минеральные вещества Наличие света и соответствующие температуры__→
→ Растительная продукция + Кислород
Не все сахара, образованные в процессе фотосинтеза, включаются в растительную биомассу, т. е. не все они идут на увеличение размеров и числа растений. Некоторая часть сахаров должна быть окислена, с тем, чтобы высвободить энергию, необходимую для биосинтеза и поддержания функций растения. В этом смысле растения не отличаются от животных: для поддержания своего существования им необходима энергия.
Можно выделить два вида продукции - общую и чистую. Общая продукция - это вся та энергия, которая была ассимилирована в процессе фотосинтеза. Чистая продукция - это накопление энергии в растительной биомассе, проявляющееся в росте или размножении растений. Разность между общей и чистой продукцией составляет то количество энергии, которое растение расходует на дыхание. Таким образом, часть общей продукции (фотосинтез) идет на дыхание (функции поддержания), а часть представляет собой чистую продукцию (рост и размножение). В большинстве случаев при исследовании продуктивности растений, особенно в наземных местообитаниях, экологи измеряют не общую, а чистую продукцию, поскольку методы ее измерения менее сложны и поскольку именно она служит мерой ресурсов, доступных гетеротрофным потребителям в данной экосистеме.
Измерение первичной продукции.
Чистую продукцию удобно выражать в граммах ассимилированного углерода, в сухом весе или в энергетическом эквиваленте сухого веса. Всеми этими показателями пользуются попеременно. Содержание энергии в каком-либо органическом соединении зависит, прежде всего, от содержания в нем углерода и азота. Содержание углерода по весу в большинстве растительных тканей близко к его содержанию в глюкозе, т. е. к 40%. Когда в растениях сахара превращаются в жиры и масла, кислород отщепляется от их молекул, так что относительное содержание углерода возрастает. Например, трипальмитинглицерид (С51Н98С6) содержит 76% углерода по весу. Содержание энергии на 1 г в жирах и маслах более чем вдвое превышает ее содержание в сахарах, а поэтому они очень часто используются растениями и животными для запасания энергии.
Соединение двуокиси углерода и воды в процессе фотосинтеза требует затраты 9,3 ккал энергии на каждый грамм ассимилированного углерода. Поэтому полное окисление какого-либо углеродсодержащего соединения до двуокиси углерода и воды должно было бы сопровождаться высвобождением ровно 9,3 ккал на 1 г окисленного углерода. Практически вследствие биохимических перестроек, с которыми связано образование большинства сложных органических соединений, количества энергии оказываются несколько иными. Принято считать, что при окислении 1 г углерода (сахаров, крахмала, целлюлозы) высвобождается 4,2 ккал энергии, 1 г белка—5,7 ккал, а 1 г жира—9,5 ккал.
Общее уравнение продукции допускает несколько возможных методов измерения первичной продукции естественных местообитаний. Поглощение двуокиси углерода и минеральных питательных веществ, выделение кислорода - все эти величины пропорциональны продукции. Для измерения поглощения двуокиси углерода и продукции кислорода и органического вещества существуют надежные методы.
Первичную продукцию наземных экосистем обычно оценивают по годовому приросту растительной биомассы (чистая продукция). Для того чтобы определить прирост за год, в конце вегетационного сезона растения срезают, высушивают и взвешивают. Этот метод сбора урожая обычно используют при определении биомассы культурных и дикорастущих растений в умеренных областях, где вся надземная часть у большинства растений ежегодно отмирает. Поскольку рост корней обычно не учитывается (из почв большинства типов корни извлекать трудно), урожай измеряет чистую годовую наземную продуктивность, которая чаще всего и используется в качестве основы для сравнения продуктивности наземных сообществ.
Метод сбора урожая сталкивается с некоторыми трудностями. Часть первичной продукции поедают растительноядные животные. Рост корней, как только что отмечалось, измерить трудно, хотя корневые системы однолетних растений иногда удается отделить от почвы путем длительного отмывания. Однако у многолетников рост корней продолжается из года в год, так что их биомасса представляет собой результат многолетнего роста. Трудности измерения, создаваемые в луговых местообитаниях ростом корней, еще больше увеличиваются в лесах вследствие роста ветвей и стволов деревьев. Сбор листового опада и обрезка свежих побегов дают возможность оценить продукцию лишь частично. Годовой прирост деревянистых частей часто вычисляют, определяя отношение окружности ствола к общей биомассе и измеряя затем годовые приросты живых деревьев. Для оценки общей биомассы срубают ряд деревьев увеличивающегося размера, отделяют стволы от ветвей, а иногда и от корней, после чего сжигают все эти части по отдельности в больших печах и взвешивают золу. Годовой прирост окружности живых деревьев можно затем перевести в увеличение общего веса. Для полной оценки продукции к полученной величине прибавляют прирост листьев, цветков и плодов, т. е. частей, которые возобновляются каждый год.
Специфической чертой продукционного процесса в водоемах служит то, что в образовании первичной продукции их большое значение имеет планктон, биомасса которого, как правило, во много раз меньше годовой продукции. Поэтому ни биомасса планктона в какой-либо момент года, например во время его максимального развития, ни средняя биомасса за год не могут служить мерой первичной продукции
В природе одновременно происходят два противоположных процесса - построение и разрушение органических веществ. Новообразование органических веществ в темноте прекращается, следовательно, прекращается и потребление углекислоты из окружающей среды и выделение в среду эквивалентного количества кислорода. Процессы же дыхания, т. е. потребление кислорода и выделение углекислоты, в темноте идут с той же скоростью, что и на свету. Поэтому, сравнив результаты жизнедеятельности водных сообществ на свету и в темноте, можно рассчитать величину первичной продукции, а в известных случаях и деструкции в водоемах.
О процессах построения и деструкции в принципе можно судить по нескольким различным показателям, например, по скорости потребления кислорода, выделения углекислоты, изменению рН, накоплению продуктов фотосинтеза и пр. В практике исследований в подавляющем большинстве случаев прибегали к измерениям содержания в воде растворенного кислорода. Следует сказать, что скорость потребления кислорода служит наиболее распространенным показателем интенсивности обмена водных организмов и скорости минерализации органических веществ, или скорости деструкции.
Наконец, биомасса водных организмов, пропорциональная количеству органических веществ, может быть выражена эквивалентным ей количеством кислорода, потребного для полного окисления. Биомасса, выраженная в единицах кислорода, оказывается непосредственно сравнимой с результатами измерений интенсивности фотосинтеза (первичной продукции) и дыхания (деструкции). Таким образом, есть основания выделить кислородный метод определения величины первичной продукции в водоемах.
При прослеживании круговорота углерода и изучении некоторых других вопросов можно было бы отдать предпочтение углекислоте в качестве единицы измерения, но для энергетического рассмотрения биотического круговорота углекислота как единица измерения непригодна, так как энергетический эквивалент ее в несравненно большей мере изменяется в зависимости от химического состава вещества. Известно, что для безазотистого дыхательного коэффициента (ДК) энергетические эквиваленты О2 и СО2 составляют следующее число калорий на 1 мл.
Таблица
ДК
|
О2
|
CO2
|
0,70 0,80
0,9
0,90
|
4,686 4,801 4,924
|
6,694 6,001 5,471
|
0,90 |
4,924 |
5,471 |
1,00
|
5,047
|
5,047
|
Для кислорода энергетический эквивалент при ДК=0,70 отклоняется всего на 7% от его значения при ДК=1,0, в то время как для углекислоты соответствующая разность составляет 33%.
Для определения величин первичной продукции планктона могут быть использованы наблюдения в дневное и ночное время в открытой воде водоемов, т. е. расчеты величин первичной продукции по суточному ходу содержания кислорода и углекислоты, или же наблюдения за результатами жизнедеятельности планктона в воде, заключенной в незатемненные и затемненные сосуды.
Наибольшее распространение получил последний способ, который может быть обозначен как метод склянок.
По истечении известного срока экспозиции, например 24 часов, в незатемненном сосуде должно содержаться больше продуктов фотосинтеза и растворенного в воде кислорода и меньше углекислоты, чем в затемненном. Очевидно, что разности в содержании кислорода или углекислоты строго пропорциональны интенсивности фотосинтеза планктона. Таким образом, об интенсивности фотосинтеза планктона можно судить, пользуясь разными критериями. Соответственно с этим можно различить по меньшей мере три варианта метода склянок. Из них первый технически наиболее простой и наиболее распространенный кислородный вариант.
Техника применения метода склянок проста. В этом существенное достоинство метода, который может быть использован, и в экспедиционных условиях. Склянки должны быть из белого стекла и иметь притертые пробки. Наиболее удобны склянки вместимостью 60— 160 мл. При таком объеме три склянки («контрольная» - для определения начального содержания кислорода, затемненная и незатемненная) могут быть заполнены из одного литрового батометра. С помощью склянок такого размера сделана большая часть наблюдений на разных водоемах, и поэтому такая вместимость их желательна для стандартизации метода.
При всех наблюдениях методом склянок, особенно при стационарных работах, рекомендуется на каждый горизонт устанавливать две светлые и две затемненные склянки, как это и практиковалось почти всеми авторами.
Когда наблюдения ведутся с помощью большого числа склянок, то при определении содержания кислорода удобно титровать не все содержимое склянки, а некоторый объем, отобранный с помощью пипетки после тщательного перемешивания раствора. При этом возможно повысить точность анализа путем повторного титрования. Например, из склянки в 120 мл можно дважды отобрать по 50 мл. При этом способе существенно облегчаются расчеты и отпадает необходимость точной калибровки склянок, но лишь в случае, когда все склянки имеют примерно один и тот же объем.
Склянки с пробами, отобранными на нужной глубине, экспонируются на той же глубине. За исключением специальных наблюдений, срок пребывания склянок в водоеме должен быть равен 24 часам. На протяжении суток начинаются и заканчиваются циклические изменения освещения и других условий.
Расчет первичной продукции по кислородной модификации метода склянок осуществляется следующим образом. Начальное содержание О2 в склянке перед экспонированием составляло V1, после экспозиции в светлой склянке содержание О2 было равно Vс, в темной склянке – Vт. Время экспозиции t = 24 ч.
Первичную продукцию (мг О2/л.ч.) вычисляют по формулам:
валовая продукция: Рвал = (Vс - Vт )/ t
чистая продукция : Рчис = (Vс – V1 )/ t
деструкция : Д = (V1 – Vт ) /t
В данном случае первичная продукция и деструкция выражены в единицах кислорода. Первичная продукция может быть выражена в разных взаимоэквивалентных единицах. При переходе от одних единиц к другим принято исходить из следующих предпосылок: оксикалорийный коэффициент для органического вещества (ОВ) смешанного состава равен 3,4 кал/мг О2; в органическом веществе содержится 46% углерода; ассимиляционный коэффициент (АК) равен 1,12, а дыхательный коэффициент (ДК) равен 0,89. При этих условиях получаем следующие переходные коэффициенты: 3,2 мгО/мгС, 10,6 кал/мгС, 0,69 мгОВ/мгО2, 2,15 мгОВ/мгС. Если АК = 1,25 при ДК = 0,80, то коэффициент перехода от мгО2 к мгС составит 0,30, от мгС к калориям – 11,3.
Радиоуглеродную модификацию метода склянок для определения первичной продукции морского планктона впервые применил Стеман-Нильсен во время работ датской морской экспедиции на судне «Галатея» (по Винберг,1960).
Сущность предложенного Стеман-Нильсеном метода сводится к следующему.
К заключенной в незатемненную склянку пробе добавляется известное количество меченой углекислоты в виде раствора карбоната или бикарбоната. Стеман-Нильсен ввел в практику исследований удобный прием предварительного приготовления ампул, содержащих нужные дозы раствора бикарбоната с меченым углеродом. Им применялись серии ампул, содержащих от 0,8 до 8 мкСu.
После некоторого срока экспозиции проба профильтровывается через мембранный фильтр. Измерив активность фильтра с осевшим на нем планктоном (г) и зная общую начальную активность внесенного радиоуглерода (R), находят отношение этих величин г/R. Принимается, что количество ассимилированной углекислоты находится в том же соотношении с общим количеством ее во взятом объеме воды (с). Определив последнюю величину гидрохимическим путем, легко рассчитать искомое количество ассимилированной углекислоты (А). Очевидно, А = r c/ R
Однако в такие расчеты приходится вносить некоторые поправки. Прежде всего допущение об одинаковой скорости ассимиляции С12 и С14 заведомо не совсем верно. Известно, что С14 ассимилируется с несколько меньшей скоростью. Это вызывает необходимость поправки на так называемую дискриминацию С14. Стеман-Нильсен считает, что меньшая скорость ассимиляции С14 компенсируется поправкой +5%.
На результаты измерений методом меченого углерода может оказать влияние также темновая фиксация углекислоты, но она обычно не превосходит 1 - 2% от скорости ассимиляции на свету. Однако в загрязненных водах, где много бактерий, она может приобретать исключительно большое значение.
Наиболее слабым местом метода является то, что с его помощью фактически измеряется не все количество ассимилированного радиоактивного углерода. Некоторая часть ассимилированных атомов во время опыта принимает участие в дыхании и вновь уходит во внешнюю среду в составе выделенных при дыхании молекул СО2. Какая доля от ассимилированных атомов С14 будет вновь отдана окружающей клетки среде, зависит от многих условий, в частности, и от продолжительности опытов. В длительных опытах вся выделяемая npи дыхании углекислота идентична ассимилированной; в этих условиях этим методом измеряется не валовая, а чистая продукция, т. е. не истинный, а видимый фотосинтез. Полагают, что при кратковременных опытах в среднем 60% выделяемой при дыхании углекислоты приходится за счет ассимилированной при фотосинтезе и что в условиях оптимального освещения интенсивность дыхания составляет 10% от интенсивности фотосинтеза. Тогда для получения истинного фотосинтеза (валовой продукции) следует внести поправку +6%. Вместе с поправками на дискриминацию С14 и на темновую фиксацию углекислоты (-1%) - это составит (+5 - 1+6) =10%. Последнюю величину (10%) Стеман-Нильсен вводит во все расчеты первичной продукции планктона. Но такая величина поправки годится только для условий, при которых дыхание составляет 10% от фотосинтеза. Когда дыхание составляет 25% от фотосинтеза, поправка равна 20%, при 50% поправка будет 50% и т.д.
Таким образом, в различных условиях поправка должна быть разной, так что любое принятое постоянное значение ее будет в значительной мере произвольным. В связи с этим Джитс предложил отказаться от внесения поправки, ограничиваясь точными указаниями, при каких условиях (сроки экспозиции, освещение и пр.) получены соответствующие данные.
Стеман-Нильсен отмечает, что при низких интенсивностях света поправка особенно велика. Поэтому он считает, что метод С14 не пригоден для измерений интенсивности фотосинтеза при очень малых интенсивностях света.
Наличие хлорофилла в клетках является необходимым условием фотосинтеза. Большое число работ показывает закономерную связь между количеством хлорофилла растительных клеток и их продукцией и биомассой. Хлорофилльный метод измерения первичной продукции фитопланктона является весьма приближенным и его использование рекомендуется только в том случае, если невозможно измерение скляночным методом. Ориентировочный расчет продукции и биомассы фитопланктона по концентрации хлорофилла "а" проводят исходя из того, что хлорофилл "а" составляет 2,5 % сухой биомассы или 6,75% содержания органического углерода. Содержание хлорофилла "а" определяют фотометрическими и спектрофотометрическими методами.
X. Одум (Н. Оdum) определял продукцию в сообществе целой реки, сравнивая изменение содержания кислорода в речной воде в дневное и ночное время и внося поправки на обмен кислородом между рекой и атмосферой. Комбинируя метод Одума с методом светлых и темных бутылей и с обычными методами сбора урожая (для тех мест, где растут крупные водоросли), можно довольно точно измерить продукцию в водных сообществах.
Для измерения фотосинтеза в наземных экосистемах удобнее использовать обмен двуокиси углерода, чем обмен кислорода, так как двуокиси углерода в атмосфере содержится меньше. Небольшие изменения содержания двуокиси углерода измерить относительно легко (в атмосфере содержится всего 0,03% СО2), и утечки из камер для взятия проб не порождают серьезных ошибок. Метод измерения продукции по двуокиси углерода сходен с методом светлых и темных склянок. Часть местообитания или даже отдельное растение заключают в воздухонепроницаемую камеру и сравнивают уменьшение содержания СО2 в течение дня с его повышением (за счет одного только дыхания) ночью. Этот метод позволяет довольно точно измерять общую продукцию.
Использование радиоактивного углерода, в частности изотопа 14С, представляет собой модификацию метода измерения продуктивности, основанную на газообмене. Когда в воздухонепроницаемую камеру добавляют известное количество радиоактивного углерода в форме СО2, растения ассимилируют радиоактивные атомы углерода в такой же пропорции, в какой они содержатся в воздухе камеры. Скорость фиксации углерода вычисляют, разделяя количество радиоактивного углерода, содержащееся в растении, на долю радиоактивной СО2 в камере в начале эксперимента. Таким образом, если растение ассимилирует 10 мг 14С за 1 ч, а доля 14СО2 в камере равна 5%, то можно вычислить, что растение ассимилирует углерод со скоростью 200 мг/ч (10:0,05). В процессе дыхания растение в конечном счете снова выделяет часть ассимилированного углерода в воздух в виде СО2 , которую оно может ассимилировать вторично. Измеряя поглощение радиоактивного углерода на протяжении 1-3 ч, можно получить надежную оценку общей продуктивности. По прошествии одного-двух дней поглощение и выделение радиоактивного углерода приближается к стационарному уровню и получаемые оценки скорее относятся к чистой, нежели к общей продукции.
Помимо С02 и воды, для синтеза органических соединений растения используют и другие вещества. Исчезновение из водных местообитаний растворенных нитратов и фосфатов может иногда служить относительной мерой чистой продукции, но только при определенных условиях: рост должен происходить быстро и растения должны превращать неорганические питательные вещества в биомассу гораздо быстрее, чем вещества эти становятся им доступными в результате разложения мертвых растений или смешивания с глубинными слоями воды. Когда процессы продукции и разложения уравновешиваются и устанавливается некий стационарный режим, то освобождение неорганических питательных веществ при разложении происходит с такой же скоростью, как их ассимиляция при фотосинтезе, а поэтому концентрация в среде растворенных питательных веществ не изменяется. Кроме того, следует иметь в виду, что накопление питательных веществ в растениях не обязательно связано какой-то постоянной зависимостью со скоростью продукции. Известно, например, что водоросли поглощают больше фосфора при высоком содержании в среде растворенных фосфатов, чем при их недостатке. А иногда растворенные минеральные вещества выделяются растениями в среду. Многие физические и химические процессы, в частности эрозия, апвеллинг и осаждение, также оказывают влияние на содержание питательных веществ в водных системах. Надежные оценки продуктивности по исчезновению из среды неорганических питательных веществ можно получить только в период «цветения» водорослей, наступающий в морях и озерах умеренной и арктической зон вслед за зимним периодом покоя. .
Сравнение нескольких методов измерения продуктивности водных экосистем было проведено на фьордовом озере Огак на Баффиновой Земле (Канада). Первичную продукцию измеряли в течение всего вегетационного сезона по поглощению радиоактивного углерода в светлых бутылях, помещенных в озеро, а кроме того, наблюдали за изменением концентрации хлорофилла, нитратов, фосфатов и растворенного кислорода (рис. 7.2). Суточная продуктивность озера быстро возрастала в начале лета, когда лед исчезал с поверхности и свет начинал проникать в глубину воды. Концентрация хлорофилла возрастала параллельно возрастанию продуктивности, а концентрации нитратов и фосфатов понижались на протяжении всего лета.
Концентрация растворенного кислорода повышалась весной вместе с вспышкой растительной продукции, но затем усиление дыхания зоопланктона затруднило выявление какой-либо прямой зависимости между концентрацией кислорода и продукцией. Любопытно, что возрастание продукции, наблюдавшееся в конце лета, не зависело ни от одного из регистрируемых факторов.

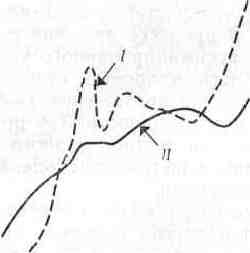
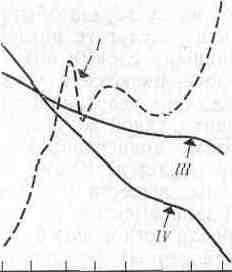
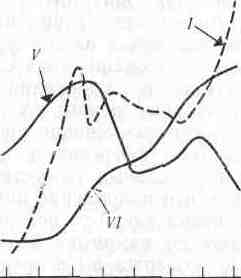
Рис. 7.2. Соотношение между продуктивностью фитопланктона (/) к концентрацией хлорофилла (II), фосфатов (х1,7) (III), нитратов (IV), кислорода (V) и зоопланктона (VI, произвольные единицы) в озере Огак на Баффиновой Земле.
Влияние факторов среды на интенсивность фотосинтеза
Свет. Изучая работу фотосинтетического аппарата растения при разных интенсивностях света, физиологи растений установили влияние света на интенсивность фотосинтеза.
При сравнительно низких освещенностях, обычно менее 25% освещенности в яркий солнечный день, интенсивность фотосинтеза прямо пропорциональна интенсивности света. Однако при более ярком свете фотосинтетические пигменты насыщаются и скорость фотосинтеза возрастает медленнее или остается на одном уровне. У многих водорослей на очень ярком свету фотосинтез замедляется, поскольку такой свет инактивирует или даже разрушает фотосинтетический аппарат.
Разные виды растений по-разному реагируют на освещенность. Так, у нескольких групп водорослей, образующих морской фитопланктон, фотосинтез достигает максимума при интенсивностях света от 0,5 до 2 ккал (м2 мин). Вместе с тем, хотя в листьях дуба и кизила световое насыщение достигается при интенсивностях, сходных с теми, при которых наступает насыщение у водорослей, интенсивности выше насыщающих не подавляют фотосинтетическую активность у этих видов. У ладанной сосны световое насыщение достигается только в самые ясные дни.
Солнечный свет, падающий на поверхность листа, состоит из лучей фазной длины волны. Не все области спектра используются в процессе фотосинтеза. Зеленые листья содержат несколько пигментов, в частности хлорофиллы и каротиноиды, которые поглощают свет и используют его энергию.
Фиолетовый Синий Зеленый Желтый Оранжевый Красный
Длина волны, нм
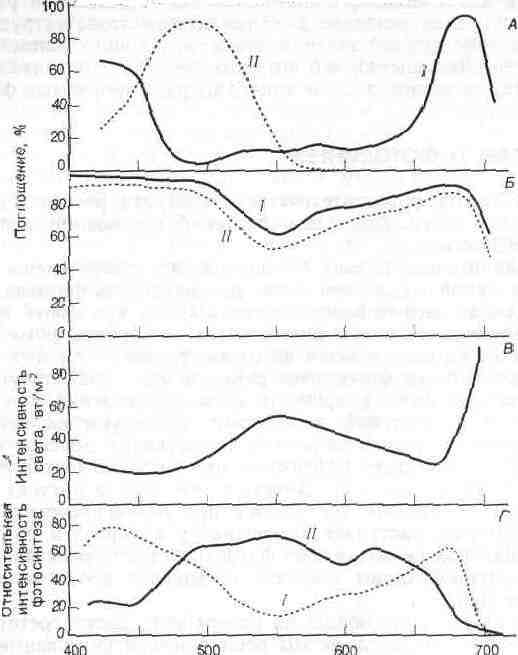
Рис. Поглощение света различной длины волны (спектр поглощения) в пределах видимой части спектра.
А. Спектры поглощения двух светочувствительных пигментов—хлорофилла (1 и каротиноидов (11). Б. Спектр поглощения целых листьев фасоли (/) и табака (//). В. Спектральное разложение света в тени леса из сахарного клена. Г. Относительная интенсивность фотосинтеза у мелководной зеленой водоросли (I) и у глубоководной красной водоросли (II) как функция области спектра. Красные водоросли фотосинтетически более активны в средней области спектра. Пигменты красных водорослей поглощают также синий и красный свет, но эти длины волн фотосинтетически относительно неактивны.
Каротиноиды поглощают свет главным образом в синей и зеленой областях спектра (рис.,А) и отражают свет с длинами волн, соответствующими желтой и оранжевой областям. Хлорофилл поглощает в красной и фиолетовой областях и отражает лучи зеленой области, которые мы и воспринимаем как цвет листьев. Спектр поглощения листьев в целом (рис. Б) примерно соответствует сумме спектров поглощения фотосинтетических пигментов, однако значительное количество света в оранжевой области поглощают, очевидно, органические соединения, не участвующие в фотосинтезе. Листья разных видов растений имеют разные спектры поглощения. Толстые, сильно пигментированные листья инжира поглощают 85% зеленого света (550 нм) — область спектра, в которой поглощение наименее интенсивно. Листья табака поглощают только 50% зеленого света. Вследствие такого избирательного поглощения света листьями свет, проникающий под полог леса, относительно богат в зеленой и инфракрасной, но беден в красно-оранжевой и голубой областях спектра (рис. В ).
Хотя вода кажется бесцветной, она поглощает небольшое количество света в красной области спектра и рассеивает голубой свет. Таким образом, вблизи нижней границы эвфотической зоны в океане преобладает зеленый свет. Фотосинтетические пигменты растений, живущих на суше и на мелководьях, слабо поглощают в зеленой области спектра, однако некоторые глубоководные морские водоросли содержат красные пигменты (эритрины), сильно поглощающие зеленый свет (рис. , Г). Эритрины в прямых лучах солнца окрашены в красный цвет, а поэтому эти водоросли называют красными. Способность красных водорослей использовать зеленый свет позволяет им проникать на большую глубину, где не могут жить зеленые водоросли, содержащие такие же пигменты, как наземные растения.
Температура. В естественных системах температура обычно тесно коррелирует с интенсивностью света, но в лабораторных условиях, регулируя эти факторы, можно изучить влияние на фотосинтез каждого из них по отдельности. При низких освещенностях, когда лимитирующим фактором является свет, изменение температуры мало влияет на фотосинтез, однако при умеренной освещенности интенсивность фотосинтеза возрастает в 2—5 раз при повышении температуры на каждые 10 °С.
Подобно большинству других физиологических функций, фотосинтез достигает максимального уровня в пределах сравнительно узкого диапазона температур, а при дальнейшем повышении температуры его интенсивность быстро падает. Поскольку листья поглощают свет, их температура к середине дня может повыситься столь значительно, что фотосинтез существенно подавляется; поэтому фотосинтез достигает максимальной интенсивности утром, около 9 часов, затем интенсивность его снижается и .вновь повышается до пиковой около 17 часов (рис.). Как и следовало ожидать, температура, оптимальная для фотосинтеза, варьирует в зависимости от среды и составляет примерно от 16 °С для многих видов умеренного пояса до 38 °С для тропических растений.
Интенсивность света, / ккал/м2.мин

Температура, °С
Рис..
А. Зависимость интенсивности фотосинтеза от температуры листьев и интенсивности падающего света. Б. Изменение интенсивности фотосинтеза в течение дня; к полудню интенсивность резко падает, так как листья слишком сильно нагреваются.
Эффективность фотосинтеза - удобный показатель для выявления скорости образования первичной продукции в растительных формациях в естественных условиях. Она выражается в процентах падающего видимого излучения, которое превращается в чистую первичную продукцию в течение тех сезонов, когда происходит активный фотосинтез. Если вода и питательные вещества имеются в достатке и не ограничивают растительную продукцию, то максимальная эффективность фотосинтеза составляет 1—2% доступной световой энергии.
Вода и эффективность транспирации. Поскольку для фотосинтеза необходим газообмен, происходящий через поверхность листа, изменение продуктивности сопровождается изменением скорости транспирации воды с этой поверхности. С понижением содержания влаги в почве извлечение из нее воды растением затрудняется и листья вынуждены закрывать устьица, чтобы уменьшить потери влаги. Когда влажность почвы снижается до точки завядания, листья надежно отгораживаются от окружающего воздуха и фотосинтез постепенно прекращается. Поэтому интенсивность фотосинтеза тесно связана со способностью растения переносить потерю воды, с наличием в почве доступной влаги и с влиянием температуры воздуха и солнечного излучения на скорость испарения. Влажная среда благоприятствует высокой интенсивности фотосинтеза, снижая транспирацию из листьев.
Агрономы используют эффективность транспирации в качестве показателя засухоустойчивости растений.
Эффективность транспирации - это отношение между чистой продукцией и транспирацией, выраженное в граммах продукции на 1000 г воды, выделенной путем транспирации. У большинства растений эффективность транспирации составляет менее 2 г продукции на 1000 г выделенной воды; у некоторых засухоустойчивых культур эффективность транспирации равна 4. Такая высокая эффективность становится возможной благодаря морфологическим адаптациям, позволяющим понизить испарение и температуру листьев. У некоторых видов, растущих в пустыне, листья по крыты волосками; эти мельчайшие волоски удерживают у поверхности листа слой неподвижного воздуха, что способствует уменьшению испарения. В холодных областях, где при замерзании почвы вода становится недоступной растениям, большинство широколиственных растений осенью сбрасывает листья, чтобы избежать высыхания, а у вечнозеленого гренландского багульника листья опушенные, и они выполняют ту же самую функцию, что и опушенные листья многих пустынных видов.
Несмотря на адаптации, благодаря которым растения снижают потери воды, они не могут избежать действия законов физики: если они вынуждены во время засухи снижать транспирацию, то они должны при этом снижать и газообмен, необходимый для фотосинтеза, следовательно, и уменьшать свою продуктивность. В некоторых тропических областях деревья во время сухого сезона даже сбрасывают листья, чтобы избежать чрезмерной потери воды.
Питательные вещества. В большинстве местообитаний при искусственном внесении удобрений наблюдается увеличение первичной продукции. Независимо от естественного плодородия местообитания внесенные питательные вещества взаимодействуют с водой, температурой и светом, определяя уровни продукции. Ограничительная роль питательных веществ, вероятно, ощущается особенно сильно в большей части водных местообитаний, в частности в открытом океане, где вследствие недостатка рас творенных минеральных веществ продукция гораздо ниже, чем в наземных местообитаниях. Обилие питательных веществ в озерах и океанах зависит от апвеллинга, глубины воды, близости береговой линии и рек, а также от характера дренирования близлежащих массивов суши. Происходящее в последнее время усиленное удобрение внутренних водоемов и прибрежных зон сточными водами и поверхностным стоком с удобряемых возделываемых земель вызвало сильное увеличение продукции в некоторых внутренних водоемах и в прибрежных частях моря. Даже кислород в определенных условиях может ограничивать продукцию наземных растений. Для роста корней и протекания в них метаболических процессов необходимо, чтобы в почве содержался кислород. В отсутствие пористости или при полном насыщении почвы стоячей водой количество доступного кислорода может опуститься ниже того уровня, который необходим для поддержания роста растений. В переувлажненных почвах болот многие растения имеют специальные приспособления, при помощи которых их корни получают кислород непосредственно из атмосферы. Например, у кипарисов имеются корневые выросты, выступающие над поверхностью воды. Эти выросты создают возможность свободного газообмена между корневой системой кипарисов и атмосферой.
Продукция в наземных экосистемах. Солнечная энергия, достигающая поверхности Земли в течение года, исчисляется величиной порядка 5-1020 ккал; в средних широтах это обеспечивает приток тепла, измеряемый 9 млрд. кал на 1 га в год.
Главной характеристикой наземной экосистемы можно считать величину продукции растительного покрова, иными словами поток энергии на уровне продуцентов.
Один гектар леса в среднем производит 6 т древесины и 4 т листвы, что при сжигании дает 27 + 19 = 46 млн. ккал. Если сопоставить полученные 9 млрд. с удержанными 46 млн. ккал, то эффективность составит около 0,5%. Следует заметить, что эти цифры определяют чистую продукцию; валовая продукция намного больше.
На рис. представлена диаграмма суммарной ассимиляции и продукции леса из обыкновенного бука.
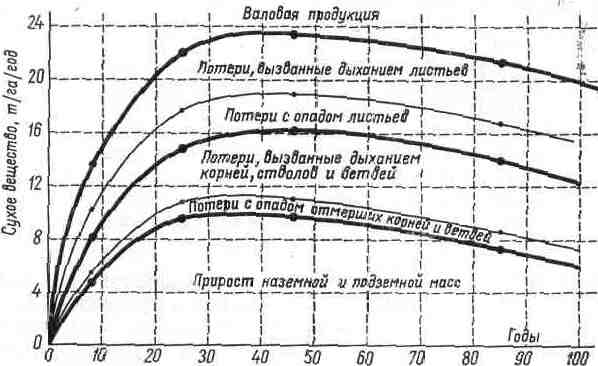
Рис.. Продукция сухого вещества в лесу из бука обыкновенного (Fagus silvatica) 2-го класса продуктивности.
Верхняя кривая представляет валовую продукцию. Жирная средняя линия характеризует чистую продукцию листвы. Пространство между названными двумя кривыми соответствует потерям при дыхании и потерям от спада. Нижняя жирная кривая — накопление древесины в надземных и подземных органах (Мо11ег, Мu11ег, Nielsen, 1954).
Кривые показывают, что в возрасте от 40 до 60 лет активность экосистемы достигает максимума: листья синтезируют 23,5 т вещества на 1 га/год. В ветви, ствол и корни направляется 16,2 т, но 40% из них в течение года составят потери на дыхание и опад отмерших частей. Из 7,3 г вещества, остающегося в листьях, 65% затрачивается на дыхание, если считать, что опад мертвой листвы составит лишь 2,5 т.
Таким образом, около половины углеводов, созданных фотосинтезом (более 10 т/га), расходуется с дыханием и, следовательно, утрачивается.
Можно заметить также, что продукция с возрастом уменьшается. Продукция надземных частей (ствол и ветви) в среднем равна 6 т/га. Вместе с продукцией корней эта средняя величина достигает 10 г/га. Если же добавить продукцию листвы, получится 12,5 т/га.
В таблице 3 представлены годовые величины продукции скольких ценных культур и нескольких естественных растительных ассоциаций.
Таблица
Чистая годовая продукция нескольких растительных ассоциаций (Lieth,1962)
|
Страна
|
Чистая продукция сухого вещества, т/га/год
|
Луг с Festuca .........
|
Германия
|
10,5—15,5
|
Луг с Lolium .........
|
»
|
5,5— 7,9
|
Луг с Arrenatherum ......
|
»
|
9,1
|
Луг с Deschampsia и клевером, 3 укоса в год . . . .... . .
|
|
|
Луг с Deschampsia и клевером, 1 укос в год .........
|
»
|
22,4 12,1
|
Болото с Sphagnum ......
|
»
|
2—10,0
|
Лес буковый .........
|
»
|
7,6—13,5
|
Лес еловый ..........
|
»
|
8,0—12,1
|
Лес из сосны обыкновенной . . . Плантация сахарного тростника . .
|
Великобритания США, Гавайи
|
6,5 34—78
|
Табл. содержит данные о культурных растениях, у которых используют лишь надземные части растений.
Приведенные показатели свидетельствуют о том, что величина продукции, равная 10 г органического вещества на 1 га в год, подтверждается для всех типов растительности. Эта величина соответствует ежегодной чистой продукции 2,75 г сухого вещества, продуцируемого на 1 м2 в день; валовую продукцию можно считать примерно вдвое большей (5,5 г).
Таблица Годовая продукция надземных частей растений нескольких растительных ассоциаций (Lieth, 1962)
Ассоциация
|
Район
|
Чистая продукция сухого вещества, т/га/год
|
Верховое болото с Phragmites . . . Верховое болото с Typha .... Низовое болото с Carex .....
|
Европа Европа, США Европа
|
2,2—25,0 10,7—17,3 4,0— 6,3
|
Заросли папоротника Pteridium . .
|
Европа
|
10,1—14,1
|
В областях умеренного климата, откуда взяты приведенные примеры, период вегетации длится не весь год, а это, естественно, снижает годовую продукцию. Продуктивность зависит и от физических условий местности, и в исключительно благоприятные годы продукция может значительно возрасти. Максимальные урожаи пшеницы, по Бойсен-Иенсену, составляют 11,9 т/га, а сахарной свеклы— 28 т/га.
В южноафриканских степях, где количество осадков изменяется от 100 до 600 мм, продукция надземных органов растений колеблется от 1 до 6 г сухого вещества на 1 га. Следовательно, за счет каждых 100 мм осадков продуцируется 1 т сухого вещества.
Таким образом, смена климатических зон обязательно сопровождается значительными изменениями продуктивности экосистем. В самом деле, продуктивность растительного покрова в альпийских и арктических зонах очень мала; продукция равняется 5 (±0,5) т/га. Напротив, она исключительно высока в экваториальных зонах, где вечнозеленая растительность функционирует все 12 месяцев в году. Приведем некоторые данные.
Саванна с преобладанием Pennisetum purpureum . 30 т/га
Плантация сахарного тростника (Гавайи) .... 67 т/га
Вторичный экваториальный лес (Янгамби) .... 20 т/га
Средние данные продукции деловой древесины (т/га/год) следующие:
Умеренные области
Лиственные 5,1
Хвойные 5,3
Тропические области
Лиственные 13,1
Хвойные 12,6
Сравнение поверхности листвы (соотношение 1:2) видов деревьев умеренных и тропических широт также показывает, что тропическая растительность на плодородных почвах и при хорошем снабжении водой в 2—2,5 раза продуктивнее, чем растительность умеренных областей. Благоприятное сочетание интенсивного солнечного света, тепла и обилия дождей делают тропики наиболее продуктивной наземной экосистемой в расчете на единицу площади. При всем этом довольно часто забывают, что в очень большой части тропиков годовая продукция значительно снижается вследствие наличия более или менее продолжительного сухого сезона.
В экосистемах умеренных и арктических областей низкие зимние температуры и длинные ночи сокращают продуктивность. Недостаток влаги ограничивает продукцию растений в засушливых областях, хотя свет и температура благоприятствуют здесь росту растений. В пределах данного широтного пояса, где количество света и температура мало изменяются в различных местностях, чистая продукция непосредственно зависит от годового количества осадков. В умеренных областях увеличение продукции начинает приостанавливаться после того, как годовое количество осадков превысит 100 см, вероятно, потому, что при этом лимитирующими факторами становятся доступность питательных веществ и свет.
Экологи Уиттэкер и Лайкенс произвели недавно оценку чистой первичной продукции для репрезентативных наземных и водных экосистем. Их оценки основаны на многочисленных исследованиях, в которых использовалось множество разнообразных методов, и, вероятно, их можно считать довольно близкими к истинным значениям.
Продуктивность наземных местообитаний резко снижается от влажных тропиков к умеренным областям; особенно резко это снижение выражено в тех местах, где климат слишком сухой или слишком холодный для того, чтобы могли расти леса. Экосистемы болот и маршей находятся на грани между наземными и водными местообитаниями, а по продуктивности растений соответствуют тропическим лесам. Максимальные уровни продукции, установленные для маршей, достигают в год 4000 г/м2 в умеренных областях и 7000 г/м2 в тропиках. Растения, обитающие на маршах, высокопродуктивны, так как их корни постоянно находятся под водой, а листья — на свету и в воздухе; тем самым эти растения одновременно пользуются благами как водной, так и наземной сред. Кроме того, они в изобилии снабжаются питательными веществами, потому что смываемый в марши детрит быстро разлагается бактериями.
Продуктивность возделываемых земель в данной области обычно несколько ниже продуктивности естественной растительности в той же .области, так как поля ежегодно перепахиваются и бывают оголены в начале и в конце вегетационного сезона, когда не нарушенные естественные местообитания продолжают создавать продукцию. Кроме того, на сельскохозяйственных угодьях обычно выращивают какой-либо один вид, который не может использовать ресурсы, предоставляемые средой, столь же эффективно, как смесь видов с различными экологическими потребностями.
Искусственное орошение и внесение удобрений могут повысить урожаи сельскохозяйственных культур вдвое и втрое по сравнению со средними величинами для всего мира. Продукция сахарного тростника, культуры, широко распространенной в тропиках, составляет в среднем около 1700 г/м2 в год. При интенсивной культуре на Гавайских островах урожаи сахарного тростника превысили средние для всего мира вдвое, а максимальная продуктивность достигла 7000 г/м2 в год. И наоборот, плохое ведение хозяйства может привести к истощению почвы и снижению продукции.
Чистая первичная продукция выращиваемых в умеренной зоне зерновых культур (пшеница, кукуруза, овес и рис), сена и картофеля колеблется от 250 до 500 г/м2год); продуктивность сахарной свеклы обычно вдвое выше. Приведем для сравнения оценки продуктивности, полученные Уиттэкером и Лайкенсом, для лесов [600—2500 г/(м2год)], и степей (150—1500 г/(м2год)] умеренной зоны. Продуктивность всех возделываемых земель, согласно данным этих авторов, варьирует от 100 до 4000 г/(м2-год), в зависимости от выращиваемой культуры, со средней 650 г/(м2-год).
Продукция в водных экосистемах. Открытый океан — это, в сущности, пустыня, где недостаток минеральных питательных веществ ограничивает продуктивность до 10% или даже менее продуктивности лесов умеренной зоны. В зонах апвеллинга (где питательные вещества выносятся на поверхность из глубины моря вертикальными течениями) и континентального шельфа (где происходит активный обмен между донными осадками и поверхностными водами) продукция выше, составляя в среднем соответственно 500 и 360 г/(м2год). Продукция мелководных эстуариев,. коралловых рифов и прибрежных зарослей водорослей приближается к продукции соседних наземных местообитаний, достигая в среднем 2000 г/(м2год). Первичная продукция пресноводных местообитаний сходна с таковой аналогичных морских местообитаний.
Различия в продуктивности водных экосистем в значительной степени определяются доступностью питательных веществ. В пределах эвфотической зоны свет, по-видимому, не является лимитирующим фактором. До 95% падающего излучения проникает сквозь поверхность воды и может быть использовано растениями. Различия в глубине, на которую проникает 'свет (зависящие от прозрачности воды), оказывают влияние на глубину, до которой может происходить фотосинтез, однако эти различия не влияют на общую продуктивность на 1 м2 поверхности океана. В прозрачных водах водоросли негусто рассеяны по всей толще эвфотической зоны, а в мутной воде они концентрируются ближе к поверхности.
Температура, очевидно, не оказывает воздействия на суммарную продуктивность морских местообитаний. У отдельных растений интенсивность фотосинтеза может быть подавлена низкими температурами, тем не менее морские водоросли достигают в холодной воде такой значительной плотности, что арктические моря столь же продуктивны, как. и теплые тропические моря. Гидробиолог К. Манн (Mann) обнаружил, что в холодных водах умеренной зоны крупные водоросли создают на 1 м2 морского местообитания такое же количество биомассы, как в Индийском океане или в Карибском море. В Новой Шотландии продуктивность только одного вида - ламинарии - достигает 1500 г/(м2.год) - продуктивность, вполне приличествующая лесу умеренной зоны!
Можно попытаться обобщить все изложенное выше и определить, согласно Одуму (Odum, 1959), мировое распределение первичной продукции, исходя из величинам валовой продукции в г на 1 м2 в день.
Выявляются величины трех порядков:
1. Очень низкой продукцией обладают океанические глубины и пустыни (0,1 г). Хотя ограничивающие факторы этих двух сред совершенно различны (в океанах - питательные вещества, в пустынях - вода), как различны и продуцирующие организмы (водоросли вместо высших растений), тем не менее и глубины океанов, и аридные территории равно следует относить к пустыням.
2. Сходной продукцией порядка 1 г (0,5—3 а) характеризуются травяные формации, морские литорали и плохо культивируемые земли.
3. Продукцией от 10 до 20 г обладают влажные леса, земли с интенсивным сельскохозяйственным использованием, аллювиальные формации, эстуарии и коралловые рифы. Максимально возможная продукция экосистемы, по-видимому, не может превышать 25 г/м2 день.
Величины продукции экосистем земного шара весьма различны и меняются в зависимости от морских или континентальных условий, от географической широты и высоты над уровнем моря, то есть от условий среды, которая оказывает значительное воздействие.
Годовая чистая первичная продукция. В наиболее продуктивных местообитаниях эффективность фотосинтеза составляет 1—2%, однако на земном шаре так много мест, в которых отсутствуют условия, оптимальные для роста растений, что лишь 0,1% всей световой энергии, падающей на поверхность Земли, ассимилируется растениями. Энергия солнечного света, падающего на внешние слои атмосферы Земли под прямым углом (солнечная постоянная), равна примерно 107 ккал/(м2.год). Однако угол, под которым падает солнечный свет, варьирует в зависимости от времени суток и времени года, и, кроме того, за год все точки на поверхности Земли находятся под покровом ночи в течение времени, эквивалентного шести месяцам. Если бы суммарную энергию, достигающую внешних слоев атмосферы, можно было равномерно распределить по поверхности Земли, то на каждый квадратный метр пришлась бы четвертая часть солнечной постоянной, или 2,5-108 ккал/(м2год). Но в действительности земная поверхность не получает так много солнечной энергии в течение года и энергия эта распределяется неравномерно. Вероятно, 40% всей световой энергии, поступающей на Землю, поглощается атмосферой и переизлучается назад в космическое пространство в виде тепла. Кроме того, часть этой энергии отражается и рассеивается частицами пыли, находящимися в атмосфере, а также поверхностями воды, горных пород и растительности.
Количество энергии, поступающей за год в определенную местность, варьирует в зависимости от широты этой местности и от облачного покрова. Местности, расположенные в умеренном поясе, обычно получают от 2-105 до 2-106 ккал/(м2 • год), что составляет от 2 до 20% солнечной постоянной. Только половина этой световой энергии может быть ассимилирована растениями; другая ее половина лежит за пределами спектра поглощения растительных пигментов.
По оценке Уиттэкера и Лайкенса, общая первичная продуктивность всего земного шара за год составляет около 730.1015ккал, из которых две трети дают наземные местообитания. Средняя продуктивность наземных областей [3200 ккал/(м2год)] соответствует ассимиляции примерно 0,3% световой энергии, достигающей поверхности Земли. Суммарная эффективность фотосинтеза в водных местообитаниях составляет менее 25% его эффективности в наземных сообществах.
Распределение продукции между основными растительными зонами земного шара зависит от локальной продуктивности этих зон, которая в свою очередь определяется температурой, количеством света, осадков и питательных веществ, а также общей занимаемой ими площадью. Тропические леса покрывают только 5% поверхности Земли, но они дают почти 28% общей продукции. Леса умеренной зоны и открытый океан, занимающие соответственно 2,4 и 63% поверхности, дают 9,2 и 25% продукции. Хотя прибрежные участки моря (эстуарии, заросли водорослей и рифы) занимают всего 0,4% поверхности Земли, они дают 2,3% ее общей продукции и 6,7% продукции водных местообитаний. Болота и марши также отличаются высокой продуктивностью в сравнении с небольшой занимаемой ими площадью.
Таким образом, продуктивность достигает наивысшего уровня в тех местах, где имеются в изобилии все необходимые для этого факторы - свет, тепло, вода и минеральные питательные вещества. Уменьшение количества света и тепла снижает продукцию местообитаний по мере их удаления от тропической зоны. В засушливых областях продукция ограничивается, кроме того, недостатком влаги. В некоторых водных местообитаниях, особенно в открытом океане, наиболее сильное ограничивающее действие на продукцию оказывает недостаток питательных веществ, но прибрежные участки, очень богатые питательными веществами, создают столько же, если не больше, органической продукции, сколько близлежащие наземные сообщества.
Чистая продукция служит мерой суммарного количества энергии, поступающего в экосистему. Эта продукция в конечном счете потребляется и рассеивается растительноядными животными, плотоядными, которые поедают растительноядных, и бесчисленным множеством организмов-разрушителей, питающихся детритом.
Общие задачи определения продукции водных животных
Процесс продуцирования в экосистеме водоема осуществляется через трофические взаимоотношения между организмами, которые заключаются в передаче определенных количеств вещества и энергии с одного трофического уровня на последующие.
Следовательно, основное значение приобретает знание тех особенностей отдельных организмов, которые определяют взаимодействие последних с организмами смежных трофических уровней, например их пищевых потребностей. При изучении закономерностей продукционного процесса биомассу, продукцию и отдельные функции организмов разных трофических уровней необходимо выражать в сравнимых единицах. Такими единицами служат единицы массы или энергии.
Выражение «единицы массы или энергии» здесь относится к миллиграммам, граммам или другим весовым единицам сухого, сырого органического вещества, органического углерода или азота, к калориям и т. д., т. е. к таким единицам, которые при известных предпосылках с большим или меньшим приближением можно с помощью некоторых коэффициентов заменить одну на другую.
В каждом отдельном случае, чтобы в конечном итоге выразить биомассу, а в особенности продукцию видовой популяции или сообществ, например в калориях, надо первоначально располагать детальными, сведениями о численности, индивидуальном весе, калорийности и о многих других особенностях и специфических свойствах изучаемых объектов.
При организации и выполнении работ по изучению продуктивности пресноводных водоемов, во-первых, сбор материала должен быть организован так, чтобы полученные данные были репрезентативны по отношению ко всему водоему; во-вторых, конечные результаты исследований должны быть представлены в виде величин, которые могут быть отнесены ко всему водоему.
Для выполнения первого требования достаточно рационально разместить станции отбора проб, места наблюдений на водоеме, горизонты взятия проб по глубинам и т. д. Чтобы это сделать, надо располагать хотя бы предварительными сведениями о характерных зонах водоема, об их относительных размерах и о главных особенностях распределения населяющих водоем организмов.
Второе требование окажется выполненным при надлежащей обработке исходных данных. В частности, при получении средних величин должны быть учтены относительные объемы слоев воды и т.д.
Только полученные таким образом данные могут служить для расчета величин, относящихся ко всему водоему.
После расчета величин, относящихся ко всему водоему, эти итоговые данные должны быть отнесены к его площади и выражены в килограммах на 1 га, граммах и калориях на 1 м2 и т. д., т. е. в форме, допускающей сопоставление с аналогичными материалами по другим водоемам.
Биомасса, т. е. вес популяции животных на определенной площади биотопа, зоны или всего водоема, в данный момент представляет собой некоторый итог противоположно направленных процессов размножения и роста с одной стороны, и элиминации - с другой. Биомасса позволяет оценить степень количественного развития видов, населяющих водоемы. Однако по этим данным ничего нельзя сказать о продукционных свойствах видов, т. е. об интенсивности роста и размножения организмов в видовых популяциях. Знание этих особенностей популяций совершенно необходимо, когда стремятся не только выявить качественные или специфические особенности трофических цепей в данном водоеме, но и количественно оценить отдельные трофические звенья и переход вещества и энергии с одного трофического уровня на последующий.
Создано большое количество крупных водоемов (водохранилищ), в формировании флоры и фауны которых наряду со стихийными процессами участвует и человек. В частности, для рационального хозяйственного использования водохранилища заселяются молодью ценных видов рыб.
Без знания скорости воспроизводства кормовой базы рыб, без определения интенсивности размножения и роста беспозвоночных, составляющих пищу рыб, невозможно установить, какую рыбную продукцию можно получить с того или иного водоема или единицы его площади. Количественные сопоставления этапов продукционного процесса в водоемах разного типа позволяют устранить субъективные оценки зависимости между кормовой базой и рыбопродуктивностью водоемов. В решении этого вопроса не последнюю роль играют методы расчета продукции водных животных.
Хотя определение продукции водных животных в первую очередь необходимо для решения вопросов продукционно-биологического направления гидробиологии, но они оказываются нужными и в санитарно-технической гидробиологии для определения функционального значения населения водоемов в самоочищении загрязненных вод. Рассматривая эти сложные процессы в самой общей форме, встречаемся, во-первых, с ролью организмов в процессе минерализации и деструкции нестабильных веществ загрязнений, во-вторых, с их стабилизацией путем превращения в живое вещество тел организмов. Первый процесс в основном пропорционален интенсивности обмена, умноженной на биомассу, второй -приросту, или продукции. Процесс самоочищения, в котором участвует многовидовое сообщество, осуществляется в значительной мере на основе трофических взаимоотношений видов. Количественно оценить эти взаимоотношения и выяснить закономерности биологического круговорота, определяющие процесс самоочищения можно только в том случае, если есть возможность судить о продукции участвующих в этом процессе видовых популяций.