

Транспорт углекислоты. Количество растворенного и связанного газа. Механизмы связывания и освобождения углекислого газа. Роль карбоангидразы и ионных транспортеров на мембранах эритроцитов.
Диффундируя в область с более низким давлением из области с более высоким парциальным давлением, углекислота переходит из крови в альвеолы и из тканей в кровь. Давление углекислоты в венозной крови - около 47, в тканях составляет около 60, а в альвеолах -около 35 мм рт. ст. В артериальной крови парциальное давление углекислоты равно около 41 мм рт.ст., поэтому кровь содержит много углекислоты и после того, как пройдет через легкие. Особым ферментом, называемым угольной ангидразой, в капиллярах тканей ускоряется примерно в 1500 раз превращение углекислоты в угольную кислоту и обратное превращение.
Газы в крови находятся в двух состояниях: физически растворенном и химически связанном. Растворение происходит в соответствии с законом Генри, согласно которому количество газа, растворенного в жидкости, прямо пропорционально парциальному давлению этого газа над жидкостью. На каждую единицу парциального давления в 100 мл крови растворяется 0,003 мл О2 или 3 мл/л крови.
Углекислый газ в крови находится в трех фракциях: физически растворенный, химически связанный в виде бикарбонатов и химически связанный с гемоглобином в виде карбогемоглобина. В венозной крови углекислого газа содержится всего 580 мл. При этом на долю физически растворенного газа приходится 25 мл, на долю карбогемоглобина - около 45 мл, на долю бикарбонатов - 510 мл (бикарбонатов плазмы - 340 мл, эритроцитов - 170 мл). В артериальной крови содержание угольной кислоты меньше. Эритроциты переносят в 3 раза больше СО2 чем плазма. Белки плазмы составляют 8 г на 100 см3 крови, гемоглобина же содержится в крови 15 г на 100 см3. Большая часть СО2 транспортируется в организме в связанном состоянии в виде гидрокарбонатов и карбаминовых соединений, что увеличивает время обмена СО2.
Карбоангидра́за фермент, катализирующий обратимую реакцию гидратации диоксида углерода: СО2 + Н2О ⇔ Н2СО3 ⇔ Н+ + НСО3. Содержится в эритроцитах, клетках слизистой оболочки желудка, коре надпочечников, почках, в незначительных количествах — в ц.н.с., поджелудочной железе и других органах. Роль К. в организме связана с поддержанием кислотно-щелочного равновесия (Кислотно-щелочное равновесие), транспортом СО2, образованием соляной кислоты слизистой оболочкой желудка.
Нарисовать пневмограмму последовательности событий: 1) спокойное дыхание, произвольная задержка и реакция на нее; 2) гипервентиляция и реакция на нее.
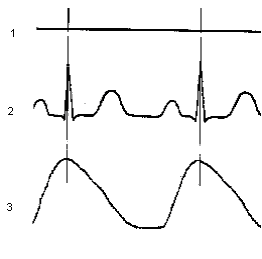
1-отметка мигания лампы фотостимулятора;
2- электрокардиограмма;
3- пневмограмма.
Вертикальные линии показывают принцип установления факта синхронизации ритма сердца и дыхания.
Билет 6
Плевральное давление. Его значение для дыхания и изменения в зависимости от фазы дыхательного цикла. Механизм нарушений дыхания при пневмотораксе.
Внутриплевральное давление — давление в герметично замкнутой плевральной полости между висцеральными и париетальными листками плевры. В норме это давление является отрицательным относительно атмосферного. Внутриплевральное давление возникает и поддерживается в результате взаимодействия грудной клетки с тканью легких за счет их эластической тяги. При этом эластическая тяга легких развивает усилие, которое всегда стремится уменьшить объем грудной клетки. В формировании конечного значения внутриплеврального давления участвуют также активные силы, развиваемые дыхательными мышцами во время дыхательных движений. Наконец, на поддержание внутриплеврального давления влияют процессы фильтрации и всасывания внутриплевральной жидкости висцеральной и париетальной плеврами. Внутриплевральное давление может быть измерено манометром, соединенным с плевральной полостью полой иглой.
Итак, дыхательный цикл включает две фазы: вдох (инспирацию) и выдох (экспирацию). Обычно вдох несколько короче выдоха: у человека их соотношение равно в среднем 1 : 1,3. Соотношение компонентов дыхательного цикла (длительность фаз, глубина дыхания, динамика давления и потоков в воздухоносных путях) характеризует так называемый паттерн дыхания.
Во время вдоха, вызванного сокращением мышц—инспираторов, вследствие увеличения объема грудной полости отрицательное давление в плевральной полости возрастает. Поэтому, а также благодаря адгезивным силам, возникающим между прилегающими друг к другу париетальным и висцеральным листками плевры, легкие растягиваются еще больше. Увеличение легочного объема, в свою очередь, ведет к падению внутрилегочного (внутриальвеолярного) давления, что и служит причиной поступления в них через дыхательные пути атмосферного воздуха.
Как только инспираторная мускулатура расслабляется, возросшая в ходе вдоха эластическая тяга легких возвращает их в исходное состояние. При этом из—за уменьшения объема легких давление в них становится положительным, воздух из альвеол устремляется через воздухоносные пути наружу.
Воздух или газ может попадать в плевральную полость снаружи (при открытом повреждении грудной клетки и сообщении с внешней средой) или из внутренних органов (например, при травматическом разрыве лёгкого при закрытой травме, либо при разрыве эмфизематозных пузырей, «булл», при минимальной травме или кашле, спонтанный пневмоторакс). В норме лёгкое расправлено за счёт того, что в плевральной полости давление отрицательное. Поэтому при попадании туда воздуха лёгкое спадается. Больной пневмотораксом испытывает резкую боль в груди, дышит часто и поверхностно, с одышкой. Ощущает «нехватку воздуха». Проявляется бледность или синюшность кожных покровов, в частности лица. Аускультативно дыхание на стороне повреждения резко ослаблено, перкуторно — звук с коробочным оттенком. Может определяться подкожная эмфизема.
Газообмен в легких и тканях. Парциальное давление газов в альвеолярном воздухе и напряжение газов в артериальной крови, венозной крови и тканевой жидкости. Особенности транспорта газов у новорожденных.
Перенос О2 из альвеолярного газа в кровь и CO2 из крови в альвеолярный газ происходит исключительно путем диффузии. Ее движущей силой служат разности (градиенты) парциальных давлений (напряжений) O2 и СО2 по обе стороны аэрогематического барьера, образованного альвеолокапиллярной мембраной. Никакого механизма активного транспорта газов здесь нет. Кислород и углекислый газ диффундируют в растворенном состоянии: все воздухоносные пути увлажнены слоем слизи. Важное значение для облегчения диффузии 02 имеет сурфактантная выстилка альвеол, так как кислород растворяется в фосфолипидах, входящих в состав сурфактантов, гораздо лучше, чем в воде. В ходе диффузии через аэрогематический барьер молекулы растворенного газа должны преодолеть: слой сурфактанта, альвеолярный эпителий, две основные мембраны, эндотелий кровеносного капилляра. Ввиду того что в транспорте дыхательных газов существенную роль играют эритроциты, к этому списку добавляются слой плазмы и мембрана эритроцита. Диффузионная способность легких для кислорода очень велика. Это обусловлено огромным числом (сотни миллионов) альвеол и большой их газообменной поверхностью (у человека она составляет около 100 м2), а также малой толщиной (порядка 1 мкм) альвеолокапиллярной мембраны. Диффузионная способность легких у человека равна примерно 25 мл О2/мин в расчете на 1 мм рт. ст. градиента парциальных давлений кислорода. При учете того, что градиент Ро2 между притекающей к легким венозной кровью и альвеолярным газом обычно превышает 50 мм рт. ст., этого оказывается вполне достаточно, чтобы за время прохождения через легочный капилляр (около 0,8 с) напряжение кислорода в ней успело уравновеситься с альвеолярным Ро2. Несколько более низкое (на 3—6 мм рт. ст.) артериальное Роз по сравнению с альвеолярным объясняется проникновением венозной крови в артериальную через невентилируемые альвеолы, а также артериовенозные шунты. Лишь при ускорении легочного кровотока, например при тяжелой мышечной работе, когда время прохождения крови через капилляры альвеол может сокращаться до 0,3 с, наблюдается недонасыщение крови кислородом в легких, что, однако, возмещается увеличением минутного объема крови. Что касается диффузии СО2 из венозной крови в альвеолы, то даже сравнительно небольшого градиента Рсо2, (6—10 мм рт. ст.) здесь оказывается вполне достаточно, так как растворимость углекислого газа в 20—25 раз больше, чем у кислорода. Поэтому после прохождения крови через легочные капилляры Рсо2 в ней оказывается почти равным альвеолярному — обычно около 40 мм рт. ст.
Рефлекторная регуляция дыхания. Взаимодействие нейронов дыхательного центра при смене фаз дыхательного цикла. Рефлексы механорецепторов легких.
Сокращения дыхательных мышц обеспечиваются ритмической активностью нейронов дыхательного центра. Неотъемлемым звеном аппарата регуляции дыхания являются также хеморецепторные и механорецепторные системы, обеспечивающие нормальную работу центрального дыхательного механизма в соответствии с потребностями организма в обмене газов. В структуру дыхательного центра входят следующие образования:
1) в двух ограниченных участках продолговатого мозга с обеих сторон располагаются инспираторные нейроны, разряжающиеся незадолго до вдоха и в течение самого вдоха (это область рострального [передне¬го, носового] отдела обоюдного ядра и область, прилегающая к одиночно¬му тракту), которые для простоты объединяют под названием "центра вдо¬ха".
2) в участке, расположенном вдоль обоюдного ядра каудальнее инс¬пираторной области, обнаружены экспираторные нейроны, разряжающиеся при выдохе и во время дыхательной паузы. Этот участок упрощенно назы¬вают "центром выдоха".
3) в медиальной инспираторной области, расположенной вдоль оди¬ночного тракта обнаружены как R-нейроны, возбуждающиеся при вдохе, так и R- нейроны, которые разряжаются не только одновременно с R-¬нейронами, но также во время паузы последних. Активность R-нейронов увеличивается при максимальном растяжении легких. Вследствие этого полагают, R-нейроны оказывают тормозное влияние на R-нейроны.
Ритмическое чередование вдоха и выдоха связано с переменными разрядами инспираторных и экспираторных нейронов. Во время активности инспираторных нейронов экспираторные клетки "молчат", и наоборот, т.е. клетки оказывают друг на друга реципрокное тормозное влияние. Для объ¬яснения механизма, ограничивающего деятельность разряда клеток, выд-вигается ряд гипотез, в одних делается упор на свойства нейронов, а в других - на тормозные межклеточные взаимодействия.
Основной регулятор активности центрального дыхательного механизма — афферентная сигнализация о газовом составе внутренней среды организма. Эта сигнализация исходит от центральных (бульбарных) и периферических (артериальных) хеморецепторов.
Бульбарные хемочувствительные зоны. На вентролатеральной поверхности продолговатого мозга расположены нейрональные структуры, чувствительные к напряжению С02 и концентрации ионов H+ во внеклеточной жидкости мозга. Локальное воздействие этих факторов вызывает увеличение дыхательного объема и легочной вентиляции. Напротив, снижение Pco2 и подщелачивание внеклеточной жидкости мозга, а также блокирование холодом или разрушение этих структур ведет к полному или частичному исчезновению реакции дыхания на избыток С02 (гиперкапнию) и ацидоз, а также к резкому угнетению инспираторной активности вплоть до остановки дыхания. Артериальные хеморецепторы. В области бифуркации сонной артерии расположен так называемый сонный, или каротидный, гломус (клубок, тельце). Он обильно снабжается кровью и содержит сложно устроенный рецепторный аппарат, реагирующий на изменения газового состава артериальной крови: повышение напряжения СО2 (гиперкапнию), увеличение концентрации ионов Н+ (ацидоз) и меньше на снижение напряжения О2 (гипоксемию). Все три фактора, вызывая возбуждение каротидных хеморецепторов, усиливают активность центрального дыхательного механизма.
Механорецепторы дыхательной системы выполняют двоякую роль: во—первых, они участвуют в регуляции параметров дыхательного цикла — глубины вдоха и его длительности; во—вторых, эти рецепторы служат источником ряда рефлексов защитного характера — кашля, например. Рецепторы растяжения легких. эти Механорецепторы оказываются источником сигнализации о растяжении дыхательных путей, а следовательно, и легких. Ирритантные рецепторы.Эти рецепторы реагируют на резкие изменения объема легких, в частности на их спадение, которое вызывает рост инспираторной активности центрального механизма, прерывая таким путем выдох. Чувствительны они и к частицам пыли, скоплению слизи некоторым химическим раздражителям. Юкстаальвеолярные (юкстакапиллярные), или J—рецепторы. Эти рецепторы чувствительны к ряду биологически активных веществ (никотину, гистамину, простагландинам и др.), проникающим либо из воздухоносных путей, либо с кровью малого круга. Рецепторы верхних дыхательных путей. Они служат главным образом источником рефлексов защитного характера, возникающих при скоплении в воздухоносных путях слизи, попадании инородных тел и химических раздражителей (кашель, глотание, чихание).
На ритм дыхания могут рефлекторно оказывать влияние раздражения различных отделов организма, а поскольку водителем ритма является ды¬хательный центр, то и афферентные пути рефлекторной дуги должны замы¬каться на дыхательном центре, а эфферентные пути идут от центра к ис¬полнительным структурам дыхательной системы. При этом можно выделить ряд рецепторных зон, оказывающих наибольшее влияние на ритм дыхания. Среди таких ВИСЦЕРО-ПУЛЬМОНАЛЬНЫХ РЕФЛЕКСОВ наиболее известны:
1. рефлекс Геринга-Брейера - если легкие сильно раздуть, то вдох рефлекторно затормозится и начинается выдох. Напротив, если сущест¬венно уменьшить объем легких, то произойдет глубокий вдох. Дуга этого рефлекса начинается от рецепторов растяжения легочной паренхимы (по¬добные рецепторы обнаружение в трахее, бронхах и бронхиолах. Некото¬рые из этих рецепторов реагируют на степень растяжения легочной тка¬ни, другие только при уменьшении или увеличении растяжения (независимо от степени). Афферентные волокна от рецепторов растяжения легких идут в составе блуждающих нервов, а эфферентное звено предс¬тавлено двигательными нервами, идущими к дыхательной мускулатуре. Фи-зиологическое значение рефлекса Геринга-Брейера состоит в ограничении дыхательных экскурсий, благодаря рефлексу достигается соответствие глубины дыхания сиюминутным условиям функционирования организма, при котором работа дыхательной системы совершается более экономично. Кро¬ме того, рефлекс препятствует перерастяжению легких.